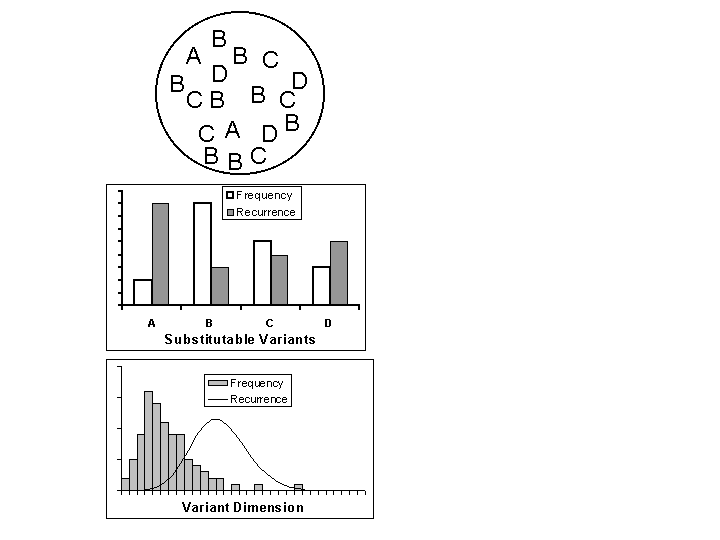Submission to Behavioral and Brain Sciences.
Draft: Do not cite or quote without the authors
permission.
(Word count = 12,529)
Mailing address:
William M. Baum
611 Mason, #504
San Francisco, CA 94108
Electronic mail: Wm.baum@unh.edu or
Short Abstract
The resemblance among operant shaping, cultural evolution, and genetic evolution appears clearly only in the light of a general concept of evolutionary process. Every evolutionary process consists of three elements: variation, recurrence, and selection. Although the metaphor of copying characterizes recurrence in genetic evolution, replication is only one type of recurrence. Selection occurs when recurrence is differential. Differences in environmental effects produce differences in recurrence, and those differences feed back to affect the composition of the pool of variants. The three evolutionary processes may be seen as nested: cultural evolution within genetic evolution, and operant evolution within cultural evolution.
(end of abstract; word count = 99)
Long Abstract
Behavior analysis is properly part of evolutionary biology, because only evolutionary theory can explain the origins of behavior and because behavior analysis follows the same mode of explanation as evolutionary theory. The resemblance among operant shaping, cultural evolution, and genetic evolution appears clearly only in the light of a general concept of evolutionary process. Every evolutionary process consists of three elements: variation, recurrence, and selection. Evolutionarily significant variation occurs among substitutable variants within a pool. These variants are defined by differences in their environmental effects. Although the metaphor of copying characterizes recurrence in genetic evolution, replication is only one type of recurrence. In cultural and operant evolution, mechanisms like stimulus control and induction cause the recurrence of the variants. Selection occurs when recurrence is differential. Differences in environmental effects produce differences in recurrence, and those differences feed back to affect the composition of the pool of variants. This general view of evolutionary process clarifies the distinction between proximate and ultimate explanations of behavior. Genetic, cultural, and operant evolution all admit of this distinction, because they all distinguish advantageous mechanisms from a history of advantage. Proximate explanations deal with the expression of variants, whereas ultimate explanations deal with the feedback from environmental effects to the frequencies of variants in the pool. The three evolutionary processes may be seen as nested: cultural evolution within genetic evolution, and operant evolution within cultural evolution. A complete understanding of human behavior requires constructing six types of explanation: proximate and ultimate explanations in all three processes.
(end of abstract; word count = 250)
Key words: evolution, general evolutionary process,
cultural evolution, operant behavior, substitutable variant, proximate
explanation, ultimate explanation, nested processes
The science of behavior, or behavior analysis, is properly part of evolutionary biology. It is intimately tied to evolutionary theory in two fundamental ways. First, evolution is the explanation of why behavior and behavioral processes exist at all. It is the only way to understand the peculiarities and constraints that characterize matters like classical and operant conditioning. Second, behavioral explanations follow the same mode of explanation as do evolutionary explanations. Skinner (1981) called it selection by consequences. It may also be called historical explanation (Baum & Heath, 1992).
Several writers have suggested that operant conditioning, or shaping, may be seen as parallel to natural selection. Donahoe (1999) suggests that even Thorndike may have recognized the resemblance between the law of effect and natural selection. Skinner (1953; 1981) stated it overtly and made it the centerpiece to his view of behavior (Ringen, 1999). Gilbert (1970) drew out the parallel at length, and Staddon & Simmelhag (1971; Staddon, 1973) enlarged on it by distinguishing between principles of variationi.e., those processes that give rise to behavioral variantsand principles of selectionthose processes that cause selection among behavioral variants. The parallel, however, is of more than casual interest; it represents a revolution in thought.
A. POPULATION THINKING
The importance of selection and history has been under-appreciated in the study of behavior within the traditions of psychology. One reason is that psychology, like other sciences, was influenced by philosophical views in which the world is thought to be composed of ideal types into which all particulars may be classified. In biology, typological thinking affected the concept of species. In psychology, typological thinking affected the concept of response, the unit of behavior. In biology, typological thinking eventually gave way to population thinking, which is the cornerstone of evolutionary theory and all selectionist theories. The difference between the two ways of thinking may be seen in their views on variation. In typological thinking, variation is error and dealt with as a nuisance to be eliminated by averaging. In population thinking, variation is central, and averaging is only an analytic convenience. As Ernst Mayr (1959) wrote, For the typologist, the type (eidos) is real and the variation an illusion, while for the populationist the type (average) is an abstraction and only the variation is real (p. 2). Arguably, Skinners greatest contribution to the study of behavior was his rejection of the typological view of behavioral units (responses). Mayr (1970) observed, The replacement of typological thinking by population thinking is perhaps the greatest conceptual revolution that has taken place in biology (p. 5). The same may be said of the (ongoing) revolution in the study of behavior.
Population thinking means the appreciation of populations as units of evolutionary change. Evolutionary theory would be impossible without it. Darwin explained the origin of species by conceiving of species as comprised of populations that change across generations. The idea of change in the composition of a population allowed evolution to be explained as descent with modification, the result of variation with consequences for reproductive success. The explanation of behavior as an outcome of selection by consequences similarly requires thinking about behavior as comprised of populations (Glenn, Ellis, & Greenspoon, 1992). Only if behavioral change is seen as change in the composition of a behavioral population, may it be explained as descent with modification. To have explanatory power, however, descent with modification or selection by consequences requires identification of an evolutionary process. Plausibility requires construction of specific explanations, which require specific mechanisms (Hull, 1988).
B. EVOLUTIONARY PROCESS
An evolutionary process includes three basic elements: variation, recurrence, and selection. Each is necessary, and together they suffice to ensure evolutionary change (i.e., descent with modification).
B.1. Variation
Variation occurs within a population or pool. A population of mice may be conceived of as a pool of genes, each mouse containing a set of genes, but because of recombination, a pool sufficiently fluid that the individual mice may often be ignored for purposes of explaining change within the population. Within the pool, different genes have different effects. Some influence coat color, some lung capacity, others the structure of the brain. The key variation within any such pool is the variation among units that affect the same trait but produce different phenotypic effectsfor example, that influence coat color but cause coat color to be dark or light. Evolutionary change depends on the existence of substitutable variants. In genetic evolution, such substitutable variants are referred to as alleles. They are substitutable in the sense that one allele may replace another, and they are mutually exclusive in the sense that such replacement is all or none. Evolutionary change consists of change in the relative frequencies of substitutable variants within the pool or population.
B.1.1. Functional definition
The units within the pool are defined, not by their structure, but by their functionthat is, by their effects. In genetic evolution, the question arises as to how to break the genome into the constituent units that influence phenotype (Dawkins, 1989a). Although DNA has structure, one cannot tell which pieces should be called genes just by examining the structure. According to Dawkins (1989b), a gene must have three properties: fidelity (faithful copying), longevity (long enough lifetime to be copied), and fecundity (frequent copying). The units that possess these properties, however, may be small or large pieces of DNA, may be contiguous in a chromosome or not, may even be in different chromosomes, and may even be in different organisms [as in Dawkinss example of parasites affecting phenotypic traits in hosts (Dawkins, 1989a and b)]. Whatever pieces of DNA act in concert to produce the phenotypic effect and may be said to be faithfully copied, to endure well enough to be copied, and to be frequently copied constitute the gene. The alleles compete because they differ in fidelity, longevity, or fecundity. A degree of copying fidelity may be assumed, because it is necessary for transmission. Differences in longevity and fecundity, however, result from differences in the alleles phenotypic effects. A darker coat color may increase its possessors likelihood of surviving long enough to reproduce.
Phenotypic effects, though often thought of in terms of morphology, are better seen as effects on the environment (Dawkins, 1989a). A darker coat color affects the ability of predators to detect a mouse against the ground. All phenotypic effects are environmental effects, because they facilitate exploitation of resources, survival, or reproduction. The point becomes clearer if we focus on genetic effects on behavior. Building a better nest alters the environment of the nestlings. Dawkins uses the example of the beavers dam construction, which creates the beaver pond, which reduces risk of predation and has a host of other good effects on the beavers environment.
This focus on function, in the form of environmental effects, solves what would otherwise be an intractable problem: how to define genes. In a broader view of selection by consequences, it solves the problem of defining the substitutable variants within the pool of variants.
The same problem of defining units arises in cultural evolution and in the shaping of operant behavior, both of which constitute evolutionary processes. As a focus on the structure of DNA offers little guidance about the definition of genes, so the structure of cultural practices and operant responses tells little about how they should be defined. Not that structure is totally irrelevant; in all three processes, structure constrains the definitions. DNA sequences specify the ordering of strings of amino acids, different configurations coding for different amino acids. Cultural practices and operant responses divide along what Skinner (1938) called natural lines of fracture (p. 33), constrained by anatomy and arrangements in the nervous system. As the codons of DNA represent natural lines of fracture and specify minimal units that may be aggregated into genes, so fixed-action patterns and the structure of bones and muscles constrain what may be aggregated into the substitutable variants of culture and operant behavior (Skinner, 1969). The making of a pot comprises motions of the fingers and hands, but only those motions that the structure of the fingers and hands allows. The physical motions, however, are little help when it comes to defining the evolutionary unit. That will be the making of a certain kind of pot, and the substitutable variants will be the making of other kinds of pot, some of which may leak less, may be easier to handle, or may serve a social function by virtue of design. These variants compete within the culture pool, just as alleles compete within the gene pool.
All three unitsgenes, cultural practices, and operant patternsare defined in terms of environmental effects. All three are defined by what they accomplish in the world with which they make contact. Dawkins (1989a) made this clear about genes in his discussion of extended phenotype. Guerin (1997) clarified the point for cultural practices when he argued that the functional unit of culture is getting a job done. Every culture comprises jobs that must get done. At the most general level, the jobs might be reproduction, obtaining resources, protection from weather and enemies, and maintaining group cohesion. These general jobs subsume more specific jobs, such as child rearing, transport, and ownership. Whatever the level of generality that suits the analysis of culture, the substitutable variants will be different ways of getting the same job done. Different ways of raising children, of transporting oneself and goods, or of demonstrating group membership may compete with one another and may differ in their outcomes. Skinner (1938; 1953; 1957) explicitly defined operant responses according to their environmental effects. One might say, following Guerin, that the rats lever press gets a job done. Although depression of a lever constitutes a discrete unit of behavior, more global patterns produce more global effects. In the laboratory, several presses may be required for reinforcement, constituting a larger unit. In the everyday world, global accomplishments always entail more specific accomplishments. Helping an unhappy customer entails listening to the complaint, making suggestions, talking to suppliers, and so on. Making a living might entail finding a job, going to work every day, specifying ones duties, and so on. Giving directions entails various utterancesstatements about location, queries, and descriptions of action. Different ways of helping the customer, of making a living, or of giving directions constitute substitutable variants that may compete and may differ in their effects. A customer may come away more satisfied, one may make a better living, directions may be given more clearly.
B.1.2. Pooling
All three processes, genetic, cultural, and operant evolution, require a pool of variation that includes substitutable variants. In general, the pool generates a frequency distributiona profile of the frequencies of various typesthat may change with time. For talking about evolutionary change, the important relative frequencies are those of the substitutable variants. Change results from competition. Figure 1 illustrates the general idea. The top diagram represents a hypothetical pool, within which are substitutable variants A, B, C, and D, which occur at different frequencies. They could represent four different alleles for four different structures of an enzyme, or four different ways to catch termites (e.g., with fingers, with a leaf, with a stick, or with a leaf stem). The relative frequencies are shown in the middle graph (open bars; the shaded bars will be discussed below). Allele (variant) B occurs at the highest frequency, then C, then D, and the least frequent is allele A. This pattern of relative frequencies may remain stable with time or may change, depending on selection.
Whereas the middle panel of Figure 1 assumes discontinuous variation, the open bars in the bottom panel illustrate a hypothetical frequency distribution for variation fine enough to be considered continuous. It could represent variation in the genes affecting height or variation in squeezing clay that affects the thickness of the walls of pots. Although the variants cannot be grouped into discrete categories, they still exhibit a pattern of frequencies that may remain stable or change, depending on selection.
Figure 1 illustrates the general idea of a pool characterized by a pattern of frequencies, which constitutes the essential element of variation in any evolutionary process. Genetic evolution, cultural evolution, and operant evolution (shaping) all assume such a pattern of variation, although they differ in details, such as whether the units are localized or extended and whether variation is continuous or discontinuous. Genes are usually thought of as particular locations on chromosomes (i.e., localized units) and alleles as differing in physical structure (i.e., varying discontinuously), but when enough genes act in concert and vary, the substitutable variation becomes (approximately) continuous. Under some circumstances, such as parthenogenesis, the unit of variation may even be the entire genome (Dawkins, 1989a). Then the substitutable variation is certain to seem continuous. Although Dawkins (1989a; 1989b) suggested a discrete unit of cultural variation analogous to the gene, the meme, nothing requires that the units of culture be localized or that variation in culture be discontinuous, any more than in genetic evolution. Boyd and Richerson (1985), for example, describe mathematical models of cultural evolution that assume variation to be continuous. In her classic book, Patterns of Culture, the anthropologist Ruth Benedict (1934) described the patterns of behavior in culture with the word custom. A custom or a practice cannot be localized to particular moments of time; it is an extended pattern of behavior that can only be observed over a substantial period. With operant behavior too, we are under no compulsion to assume discontinuous variation or localized units. That variation may be continuous is recognized in the study of response dimensions such as force, duration, and rate. The assumption that behavior must be defined in terms of momentary response units, however, has persisted for over a hundred years. Nineteenth-century connectionism, in the forms of associationism and reflexology, promoted a view of behavior as composed of localized units such as ideas, sensations, and responses. That view was an accident of history, however, and may be overcome. Baum and Rachlin (1969), for example, suggested that behavior be divided into periods filled with various activities. Favorite responses of the laboratory, such as the lever press and the key peck may be recast as activities, lever-pressing and key-pecking, which extend in time (Baum, 1976). Rachlin (1994) suggested that behavior generally be thought of as organized into patterns extended in time (Baum, 1995a, 1997). As with the customs of culture, an individuals daily behavior may be divided into activities like working, attending to family, and entertainment, none of which may be localized to moments in time.
Frequency distributions like those in Figure 1 presuppose that one may characterize the variation in a gene pool or behavior pool by taking a sort of snapshot of the frequencies at a moment in time. This might seem necessary for analytical purposes, because the frequencies may be changing. In practice, however, the snapshot is an abstraction, just as instantaneous velocity in physics is an abstraction, because any real sample is extended in time. In a gene pool, birth and death constantly alter the picture. For a behavioral pool, temporal extension is unavoidable. The customs of a culture or the habits of an individual cannot occur at a moment in time. The problem of taking a snapshot of frequencies is solved by choosing a time period long enough to provide an adequate sample but short enough that the pool may be thought of as unchanging for that duration. Depending on rate of change, one might gather data over a period of weeks, months, or years. As long as change in the pattern of frequencies during a period of sampling is negligible in comparison with change between samples, the course of change in the pool may be studied. That population thinking is central to genetic evolution has long been recognized. Population thinking, however, is equally central to understanding evolution of cultural behavior and individual operant behavior (Glenn, Ellis, & Greenspoon, 1992).
B.1.3. Cultural practices and operant behavior
The line between cultural practices and operant behavior patterns may be fine. Indeed, Skinner (1981) argued that cultural patterns are operant patterns. Since cultural practices presumably are maintained by their consequences, they might be considered operant patterns by definition. They may be distinguished, however, for the purpose of discussing cultural evolution by two characteristics. First, cultural practices are the possession of a group. To be called a cultural practice, a behavior pattern must occur in the members of a grouppossibly a subgroup within a larger group, but definitely in more than one individual. Second, cultural practices are transmitted from member to member in the group. Unlike individual operant patterns, which one may say are transmitted to the same individual at different times, cultural practices are transmitted from one individual to another and may occur in different individuals at the same time.
B.2. Recurrence
Recurrence means occurring again or coming up again. It is a general term for the tendency of a type of unit to reappear, with variation, time after time in the populationa more general term than replication, transmission, or retention (Campbell, 1965).
B.2.1. The copying metaphor
Genetic evolution is usually taken as the paradigmatic example of an evolutionary process. As an example, genetic evolution has both advantages and disadvantages. An advantage is that it has received enough attention to be both familiar and relatively well understood. A disadvantage is that, being comparatively well understood, some of its peculiarities are too easily mistaken for properties of evolutionary processes in general. This is particularly true of its mode of recurrence, which is often described with the metaphor of copying.
Evolutionary recurrence need little resemble copying a page in a photocopier. The word replication may be a synonym for copying, but may also be equated to reconstruction, which, for DNA, would be closer to the mark. We know about the uncoiling of the strands and the assembling of amino acids into replicate strands, with some recombination and occasional errors. In meiosis, the entire genome is replicated and divided into haploid components at once. The entire haploid genome is passed as a whole to the next generation. We need to ask, however, which properties of genetic transmission are essential for evolution. Darwin knew nothing of what we know about genetics today. Yet he was the author of the theory of evolution. His ignorance of genetics in no way prevented him from seeing the role of recurrence.
Darwin understood what was necessary: that the traits of parents tend to recur in offspring. More generally, the traits of one generation are passed on to the next generation. Even the words parent, offspring, and generation, impede generalization. The essential part is recurrence through time. Traits tend to breed true. In the gene pool of a population of mice, the genes that make for a dark coat tend to recur through time, with the result that dark coats tend to recur (and possibly lower predation rate). In the beavers gene pool, the genes that make for dam constructing tend to recur, and if substitutable variants exist, each variant tends to recur, although selection may favor one variant over others. True, the recurrence of genes is the result of the details of reproduction, and those details are essential to understanding many aspects of genetic evolution. For a general definition of evolutionary process, however, what counts is the tendency of types in a population to recur in time.
Today we forgive Darwin for having believed in the inheritance of acquired characteristics, but writers about culture point out that such inheritance does occur in cultural evolution (e.g., Boyd & Richerson, 1985). Again, however, the mechanism of recurrence is only important for understanding the details of cultural evolution. That it entails the inheritance of acquired characteristics helps us to understand the ways in which cultural evolution differs from genetic evolution. It is faster, for example, because transmission may occur between any individuals, not just parents and offspring, and may occur throughout the lifetime of an individual. It occurs as a result of imitation and instruction, although we might argue about exactly what those processes entail (Baum, 1994; 2000). The key, however, is that if a potter makes a certain kind of pot, his students will make the same kind of pot, and their students also, and so on. The practice recurs through time in the cultures pool of practices.
Suppose, instead of genetic evolution, we took operant evolution (i.e., shaping) as the paradigmatic example of an evolutionary process. We would see immediately that copying or replication is just one means of recurrence. A pigeon in an experiment pecks at a key time after time. A person goes to work day after day. Too little is known about the nervous system to say what the mechanism is by which behavior recurs in the same organism from time to time. Luckily, we may proceed with studying behavior without having any idea how recurrence is accomplished. The situation resembles that in biology prior to the synthesis of genetics with evolution in the early part of the twentieth century.
B.2.2. Imperfect recurrence
Part of the reason for variation is that recurrence is often imperfect. Hull (1988) defines a replicator as an entity that passes on its structure largely intact in successive replications (p. 408). A paraphrase for the sake of generality might substitute recurrences for replications. Emphasis, however, should fall on the word largely. Accidents happen. In genetic evolution, reconstruction of DNA sometimes goes awry. In cultural evolution, imitation and instruction may be inexact. In operant evolution, too, variation is intrinsic to recurrence, because context never exactly repeats, reconstruction even of a stereotyped activity like key-pecking varies, and accidents happen. Recurrence need only have high fidelity; it need not be perfect. Indeed, one might argue that it must be imperfect, if the pool is to include sufficient variation to produce novelty. Without novel types, a pools response to selection must eventually cease.
B.2.3. Recurrence in behavior
In genetic and cultural evolution, the units (genes or practices) may outlive their possessors [their vehicles in Dawkinss (1989b) parlance; their interactors in Hulls (1988)]. Operant behavior, in contrast, must die with its possessorby definition, because if it lived on in others, it would be considered cultural. In no way, however, does this disqualify operant shaping as an evolutionary process. The key elements remain: a pool of variation, substitutable variants, and recurrence of variants through time. The frequency distributions in Figure 1 might describe genetic variation, cultural variation, or operant variation. Whether they remain stable or change with time depends on their relation to selection.
B.3. Selection
The key to selection is differential recurrence. If substitutable variant A1 tends, on the average and over time, to recur more frequently than A2, the relative frequency of A1 increases at the expense of the relative frequency of A2. This happens if the size of the pool of variants is fixed or increases more slowly than the variants rate of recurrence. Usually one assumes the pool size to be fixed, because the size of a population of mice, for example, is limited by the environments carrying capacitythat is, the resources available to support the population. A behavioral pool, of cultural variants or operant variants, is fixed by limits on time, because only so much behavior can occur in a 24-hour day. Consequently, when one substitutable variant increases in frequency, another must decrease. If one variant is more successful, it tends to replace the other.
One substitutable variant succeeds over another by virtue of superior environmental effects. For genetic evolution, this is where longevity and fecundity come in. Genes endure if their vehicles (interactors) endure, and recur if their vehicles (interactors) reproduce. The specific reasons for enhanced recurrence may be extremely variedbetter defending or capturing of resources, better avoidance of predators, better defense against parasites, better mate selection, more mates, more offspring, better care of offspring, and so on. All of these superior environmental effects ultimately increase the recurrence of the alleles that produce them and thus decrease the recurrence of the less successful alleles. For cultural and operant evolution, the same sorts of environmental effects act on the pool, but less directly and, therefore, with some slippage.
B.3.1. Selection in culture
The consequences of cultural practices that may vary, making one variant more successful than another, may be thought of in terms of reinforcement and punishment. Practices are maintained proximally by social reinforcement and punishment (Baum, 1995b, 2000; Skinner, 1981). Our group shuns the eating of pork is backed up with punishment (e.g., disapproval, ostracism) for eating pork. This mode of adornment is correct is backed up with reinforcement (e.g., status, mating opportunities) for adorning oneself so. Ultimately, the maintaining and differentiating social reinforcement and punishment are cashed out in terms of reproductive success of the vehicles (interactors) that carry the genes producing susceptibility to social consequences (docility; Simon, 1990). If group membership is beneficial to reproductive success, then group cohesion and cooperation are beneficial, and practices that make for group cohesion and cooperation are selected as a result of the members docilitytheir susceptibility to social consequences. Indeed, Boyd and Richerson (1992) demonstrated theoretically that social punishment allows selection of cooperation and just about any other behavior salutary to the group. [This summary omits other adaptations, such as sensory specializations and imitation (Baum, 1994; 2000).]
Dawkins (1989b) argued that the slippage between proximal social consequences and ultimate reproductive consequences allows cultural evolution to proceed independently of genetic evolution. The logic of genetic evolution goes against such a view. If some alleles make for more docility than others, the most successful alleles lie somewhere in the middle of the range of possible levels of docility, somewhere in between none and anything goes. Too much docility means too little certainty of reproduction, because too much docility allows the spread and maintenance of customs that reduce reproduction. Thus, ultimately, alleles producing too much docility are selected against. [Boyd and Richerson (1985) make a similar point in their discussion of the tension between imitation and individual learning.]
One factor that curbs docility is primary reinforcement and punishment. As Dawkins (1989b; p. 57) suggests, consequences like sweet taste in the mouth, orgasm, mild temperature, smiling child or various sorts of pain, nausea, empty stomach, screaming child may have a genetic base because their presence generally affects the likelihood of successful reproduction. Such stimuli constitute proximate tokens of ultimate reproductive success or failure. If an interactor (vehicle) could be put together that would increase any behavior that produced the nice environmental effects (circumstances that enhance reproductive success) and avoided the nasty environmental effects (circumstances that depress reproductive success), such an interactor (vehicle) would prosper, and genes it carried would be more likely to recur in subsequent generations. As a result, genes that make for susceptibility to reinforcement by fitness-enhancing tokens are selected. As a result, neither individual behavior nor cultural customs will stray too far from patterns that maintain the frequency of those tokens. They will stray, however, for two reasons. First, the tokens bearing on reproductive success is far from certain. Sweet taste is a reinforcer, and some consumption causes no problems, but over-indulgence in sweets undermines health. Second, tokens may conflict with one another. The same behavior that produces a smiling child may also produce an empty stomach. Which should win out may be far from clear; only the calculus of long-term reproduction can tell. When short-term and long-term consequences conflict this way, dysfunctional behavior may arise. Whether it persists or not depends on whether patterns that enhance fitness in the long run, such as eating fruits and vegetables, replace the dysfunctional ones, such as eating junk food (Baum, 1994, 2000; Rachlin, 1995). Although a person may die before dysfunctional behavior is replaced, correction in culture lies under no such limitation. It may take a few generations.
B.3.2. Primacy of reproductive success
Maladaptive customs tend to drop out of the culture pool for two reasons. First, failure to correct results in natural selection. Those who over-indulge in sweets tend to become sick with diabetes and leave fewer offspring as a result (Diamond, 1992). Maladaptive customs decrease in frequency when their vehicles (interactors) leave fewer descendants to continue the customs. Groups that follow dangerous or abstinent practices, such as Quantrills Raiders or the Shakers, tend to disappear. Second, but probably more important, is that maladaptive customs contrast with their competitors (substitutable variants) in the calculus of reinforcement and punishmenti.e., in the tokens of fitness. Prohibition in the U. S. was a response to the ruinous effects of over-indulgence in alcohol; today educational campaigns encourage patterns of moderation or abstinence, based on better quality of life. When a maladaptive custom like smoking tobacco spreads through a culture because of short-term reinforcement, its long-term punishing effects eventually come into focus in the form of social contingencies that punish it more immediately (or reinforce alternatives like nicotine patches more immediately).
The power of reinforcers and punishers as tokens of long-term reproductive success makes persistence of maladaptive customs unlikely. The genetic underpinnings of reinforcement and punishment argue against Dawkinss (1989a and b) conjecture that cultural evolution proceeds independently of genetic evolution. The genes that underlie learning and culture open the door, so to speak, to environmental influence, but they do not fling it wide.
Selection in all three processesgenetic, cultural, and operant evolutionmay be traced back to reproductive success of genetic variants. In genetic evolution, the relation is direct; in cultural and operant evolution, the relation is less direct, but still strong. When genes that make behavior sensitive to its consequences are selected, that must mean that the advantages of behavioral flexibility outweigh the costs it imposes in terms of energy and risk of error. If not, competitor genes that allowed no flexibility would prevail. So, tokens of reproductive success attained the status of reinforcers, because variation in the environment rewarded flexibility in the means to them. Signs of health, resources, and social relationships became powerful reinforcers, because the interactors (vehicles) for which they were reinforcers reproduced more often. A parallel argument applies to punishers, such as nausea, pain, snakes, and frowns. The same mechanisms of reinforcement and punishment that select among substitutable variants in operant evolution select also in cultural evolution. The differences are that cultural evolution entails transmission from individual to individualimitation and instructionand therefore depends heavily on social stimuli for provenance and social reinforcement and punishment for selection.
In all three of the processes we are discussing, a question arises about the extent of the vehicles or interactors. Hull (1988) defines an interactor as an entity that interacts as a cohesive whole with its environment in such a way that this interaction causes replication [i.e., recurrence] to be differential (p. 408; italics in the original; bracketed material added). But how large a unit can interact as a cohesive whole? Group selection is usually treated with skepticism among evolutionary biologists, because it should normally be too weak to have much effect on gene frequencies. To work, it would require that gene flow into the group be negligible. That might be true of colonies of eusocial insects, for example (Seeley, 1989), but it would be unlikely for most groups, because of immigration of new members into the group. In cultural evolution, it is more likely, because imitation and instruction maintain a high degree of conformity within the group, even if new members join (Richerson & Boyd, 1992). In cultural group selection, groups with certain practices (e.g., a type of food cultivation) may out-compete other groups. The advantaged group may increase in frequency if it reproduces by fission and if competing groups tend to dissolve. In evolution of individual operant behavior, where the distinction between replicators and interactors disappears, the analog to group selection is the temporally extended contingency. Extended contingencies are often discussed in terms of delays of reinforcement. The more temporally extended the behavioral pattern, the more delayed the consequences. Alternatively, one may consider delay to be incidental and the crucial aspect of extended patterns to be their cohesiveness. Extended behavioral patterns, such as eating a good diet, are notoriously difficult to maintain. Rachlin (1995) argues that good extended patterns (often called self-control) have a greater payoff than short-term patterns (called impulsiveness or defections from the larger pattern). A defection from eating a good dieteating an ice cream sundaehas an immediate payoff, but a lower value than eating a good diet in the long run. Selection on extended patterns is weak for the same reason that group selection on genes is weak. The boundaries of the group or of the extended pattern tend to be permeableto immigration or to defectionthe greater the permeability, the weaker is selection. This may be overcome in operant evolution by introducing relatively short-term contingencies that maintain the extended pattern intact (e.g., reminders about ones diet).
B.3.3. Why not phenotypic plasticity?
To explain the variation of culture or behavior from one environment to another, the alternative to selection is phenotypic plasticity. Cosmides and Tooby (1992; Tooby & Cosmides, 1992), for example, put forward the idea that the human brain contains a large number of modules, algorithms, or mechanisms that produce behavior depending on environmental conditions. They contend, If human thought falls into recurrent patterns from place to place and from time to time, this is because it is the expression of, and anchored in, universal psychological mechanisms (p. 216). They refer to such patterns as evoked culture. The conception has been compared to a jukebox containing recordings that can be played whenever called upon (Wilson, 1999). When food availability is highly variable, the social contract algorithm plays out food sharing; when food availability is stable, the jukebox plays keeping food within the family. Such explanations fail on two grounds. First, they are implausible. They exaggerate what is probably true, that genes constrain evolution of cultural practices and of individual behavior, to make a claim that would skip over the obvious effects of consequences on behavior. Instead of sensitivity to feedback, they substitute input-output rules; given a certain environmental input, the mechanism produces a certain output. Such a view fails to explain an obvious fact: that culture evolves even if the environment remains constant. Second, alternative explanations are more plausible. If human behavior falls into recurrent patterns, that may be because of convergent cultural or operant evolution. Similar contingencies select similar behavior.
C. EVOLUTIONARY EXPLANATIONS OF BEHAVIOR
Mayr (1961) distinguished between proximate and ultimate explanations of behavior. Alcock (1993) incorporated the distinction into a textbook on animal behavior. According to Alcock, proximate explanations explain how mechanisms within an animal operate, enabling the creature to behave in a certain way (p. 2; emphasis in the original). Ultimate explanations explain how those mechanisms evolved as a result of selection. Proximate explanations refer to physiology and development. Ultimate explanations refer to history and reproductive success.
C.1. Genetic proximate and ultimate explanations
Proximate explanations are about individual organisms, whereas ultimate explanations are about populations. The question, Why do beavers build dams? may be answered in two different ways. In one interpretation, the account would refer to stimuli from water and trees, the structure of the nervous system, hormones, and so on. It would explain how the cluster of genes that make for dam building express themselves in physiology and interact with the environment to ensure that Beaver X, alive today, builds a dam. That would be a proximate explanation. The ultimate explanation, in contrast, would make no reference to Beaver X, but would be about beavers as a species or about a population of beavers or about the gene pool of a population of beavers, to which Beaver X or its genes might belong. It would refer to the adaptive consequences of building dams, variation within populations long ago, and increase in the frequency of alleles promoting dam building. It would explain how those alleles became common in the gene pool. Proximate explanations are silent about where the mechanisms for dam building came from. Ultimate explanations are silent about Beaver X, except to say that Beaver X builds a dam because it is the nature of beavers to build dams, and then to explain where that nature came from. So to speak, ultimate explanations are about the forest, whereas proximate explanations are about the trees.
Proximate explanations explain the behavior of individual organisms in terms of present mechanisms, whereas ultimate explanations explain patterns of frequency within a pool, such as those shown in Figure 1, in terms of a process of differential success. Proximate explanations refer to causes in the present, whereas ultimate explanations rely on processes that may be called historical, because they extend in time (Baum & Heath, 1992). Evolutionary change results from continual operation of the process of selection over a period of time. That it takes time may be incidental, because it is a process of adjustment with a beginning in a disturbance (in the environment or in the population) and an end in a stabilized population. It is understood as a whole; at any point in-between, it is incomplete. Suppose the shaded bars in the middle panel of Figure 1 represent the relative rates of success (recurrence) from one time period (e.g., generation) to the next. Although variant A is lowest in frequency, it has the highest rate of recurrence. Given this pattern, we expect the frequency of variant A to increase, and the frequencies of B, C, and D to decrease. If relative recurrence is independent of frequency, the process will be incomplete until A dominates. (If relative recurrence is frequency dependent, a different equilibrium results, but that in no way affects this discussion.) Suppose the curve in the lower panel of Figure 1 shows relative recurrence for the continuous case. The situation is essentially the same; we expect the frequency distribution to shift to the right. Eventually, we expect its maximum to coincide with the maximum of the curve; then the distribution stabilizes, and selection acts to keep it stable.
C.2. Cultural proximate and ultimate explanations
Since every evolutionary process includes the distinction between the mechanism that produces the advantageous behavior and the history of advantage for that behavior, cultural evolution and operant evolution also admit of the distinction between proximate and ultimate explanations (Alessi, 1992). In cultural evolution, the mechanism by which a custom is transmitted explains why it persists (i.e., why it recurs), whereas the prevalence of the custom ultimately lies in a history of competition and selection. As with the question, Why do beavers build dams? the question, Why does this tribe adorn itself with tattoos? may be interpreted and answered in two different ways. In the proximate interpretation, the question might be reworded as, Why do the members of the Hells Angels motorcycle club wear the clubs tattoos? Alternatively, Why does Tom, who belongs to Hells Angels, wear the tattoos? Three or more mechanisms of transmission might be involved. Tom might imitate other members of the tribe. Other members might instruct Tom: If you want to be accepted, you should get the tattoos. Once Tom began getting tattoos, reinforcement from tribe members might lead to getting more tattoos. In the ultimate interpretation, the question might be reworded as, How did it come about that the Hells Angels wear those tattoos? The answer would refer to the history of selection in the culture pool. Tattooing may have competed with other forms of adornment, such as wearing black jackets, wearing hair in a ponytail, or speaking in a certain dialect. One or two members may have gotten tattoos. Other members who saw them frequently may have imitated them as a result. Boyd and Richerson (1985) call this frequency-dependent bias. The first members with tattoos may have been imitated by the other members or may have been able to instruct the other members because they held high status in the club. Boyd and Richerson (1985) call this tendency to imitate success indirect bias. Tattooing may have worked better to identify members because other groups might wear black jackets or ponytails and because dialects vary from region to region; tattoos are permanent and unambiguous. Boyd and Richerson (1985) would call reinforcement of a custom by such an environmental effect direct bias. Any combination of indirect bias, frequency-dependent bias, and direct bias would result in an increase in the frequency of wearing tattoos among the tribe, until the wearing of tattoos became virtually universal.
C.3. Nesting of evolutionary processes
Genetic evolution may be thought of as an overlay on cultural evolution. Ultimate and proximate genetic explanations may be constructed for the question, Why do the Hells Angels wear tattoos? They would be analogous to the explanations of dam building in beavers. The proximate rewording of the question would be, Why did Tom get himself tattooed? As with Beaver X, the explanation refers to stimuli (e.g., from the behavior of other members), reinforcers (e.g., from the other members), development (e.g., early exposure to Hells Angels), and physiology (e.g., brain mechanisms)all of which might be traced to gene expression. The genetic ultimate explanation rewords the question, How is the wearing of tattoos beneficial to the members of the tribe? The answer might be that it constitutes symbolic marking of the group, distinguishing it and promoting cohesion within it, which in turn promotes group selection for various forms of cooperation, which enhance the success of the group and thus boost the members reproductive success. It would include also the advantages to alleles that make for frequency-dependent bias, indirect bias, and direct bias in competition with alleles that do not (Boyd & Richerson, 1985; Richerson & Boyd, 2000). Other stories might be told, but they would all end with the same reference to reproductive success, because in a genetic ultimate explanation genes must be selected. The biasing effects of genes ensure that any custom that decreases reproductive success, even if common for a time, is likely to disappear eventually.
C.4. The general evolutionary process
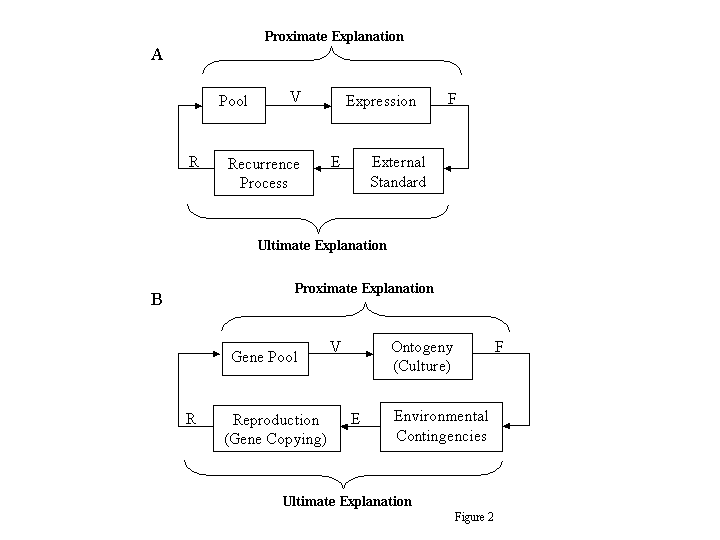
Figure 2A diagrams an evolutionary process in general terms and shows the different focuses of proximate and ultimate explanations. Each of the variables, V, F, E, or R, may be thought of either as a set of variants, as a frequency distribution across variants, or as a frequency of a particular variant relative to all others. In the parlance of cybernetics, they are operands and transforms, whereas the rectangles represent processes (transformations) that produce transforms from operands (Ashby, 1956). A pool of variants (process) results in V, a distribution of substitutable variants or relative frequency of a particular variant relative to all its competitors. A process of expression transforms V into F, a distribution of interactors (or vehicles) or frequency of one type of interactor relative to all others. The expression process receives input from the environment, omitted from Figure 2 in the interest of simplicity. The input is represented by levels of variable N (N1 N6) in Figure 3, which illustrates the various transformations in Figure 2A. The top line of Figure 3 shows Vi composed of three variants, A, B, and C, having equal frequencies. The different levels of the environmental variable N affect the outcomes in Fi; so that B coupled with N2 goes to a, the same outcome as A coupled with N1, whereas B coupled with N3 goes to b, and C transforms to b, g, or d, depending on N. In this illustration, the variation in N results in a distribution Fi that has more variety than Vi.
A set of external standards or contingencies, which might be characterized as a set of if-then rules, transforms F into E, a distribution of external effects. These are shown in Figure 3 as levels of W, X, Y, and Z, just to emphasize that they are transforms of the elements of Fi: W goes with a, X with b, Y with g, and Z with d. They may be thought of as levels of successe.g., reinforcer rates or numbers of matings. E feeds back to a recurrence process, which results in Ri, a distribution of recurrence rates or a relative recurrence rate. Following the example in Figure 3, if the elements of E associated with A (W1 W4) have a collective weight of 10 (6 + 3 + 1), those associated with B have a collective weight of 6, and those associated with C have a collective weight of 8 (entirely due to the fortunate occurrence of N6, which resulted in transform d in Fi), then the frequencies in Ri reflect these collective success rates. The appearance of variant D, however, has nothing to do with these success rates, but rather represents some kind of accident: a mutation, copying error, or external force. Ri closes the loop by entering the pooling process to result in the new distribution of variants Vi+1.
If this system is disturbed by a change in the pool (e.g., mutation or immigration) or in the external standards (e.g., a change in climate), resulting in disequilibrium like that depicted in Figure 1, it will tend to move back toward equilibrium, because iterations of the feedback loop cause the composition of the pool to change until it comes into accord with the external standardsi.e., to an optimal fit.
Figure 2A also illuminates the roles of proximate and ultimate explanations (braces). The links from the pool to the external standardsi.e., from V to Fconstitute the focus of proximate explanations. They pose the question, By what mechanisms does a distribution of variants result in a distribution of interactors? The links from the external standards to the pool constitute the focus of ultimate explanations. They pose the question, How does the distribution of external effects change the composition of the pool? That, of course, is a question about selection. The diagram omits any explicit indication of the iterative nature of evolutionary change; that must be taken for granted. The reason that both proximate and ultimate explanations are necessary for a full understanding is that the two explanations address different parts of a whole processmechanical connection and feedback, immediate causation and history.
C.5. Genetic evolutionary process
Figure 2B shows the general diagram applied to genetic evolution. The gene pool offers a distribution (V) of genotypes. Ontogeny or development transforms V into a distribution (F) of phenotypes. Environmental contingencies specify a distribution (E) of environmental effects, such as rates of obtaining resources, of predation, or of finding mates. Reproduction, which may be thought of as gene copying, depends on E. Sometimes it is thought of as simple transmission of genes from parents to offspring, but it may also be complex, because it may be the locus of other forms of selection besides natural selectione.g., sexual selection. Its outcome R, the distribution of copying rates (fitnesses), in turn changes or maintains the composition of the gene pool. Proximate explanations focus on processes of ontogeny and development. Ultimate explanations focus on the feedback, the way the environmental contingencies and reproduction achieve selection among genotypes.
As an example, we may apply Figure 2B to the evolution of culturethat is, the transition from a non-cultural species to a cultural species as a result of genetic evolution (as opposed to cultural evolution, our second process). Ontogeny converted the distribution of genotypes V into a distribution of phenotypes (F) that varied in ability and propensity for culturethat is, in the mechanisms that cause a group of people to have a culture (group-level behavior transmitted from member to member). Elsewhere I have argued that three mechanisms would suffice: sensory specializations, imitation, and social reinforcers (Baum, 1994, 2000). Whether or not these are the answer, they suggest the sort of mechanisms that would constitute a proximate explanation for the development of culture in children and newcomers to a group. The distribution F might be called variations on culture. A highly variable environment made culture beneficial to the hominids that possessed even its rudiments (Boyd & Richerson, 1985; Richerson & Boyd, 2000). It would, for example, solve many problems related to the obtaining of resources and mates. Those variants favored in the distribution of environmental effects (E) would be more represented in distribution R, with the result that the genes underlying the mechanisms for culture would increase in the gene pool. That feedback would be the ultimate explanation for the existence of culture.
C.6. Nesting cultural evolution within genetic evolution
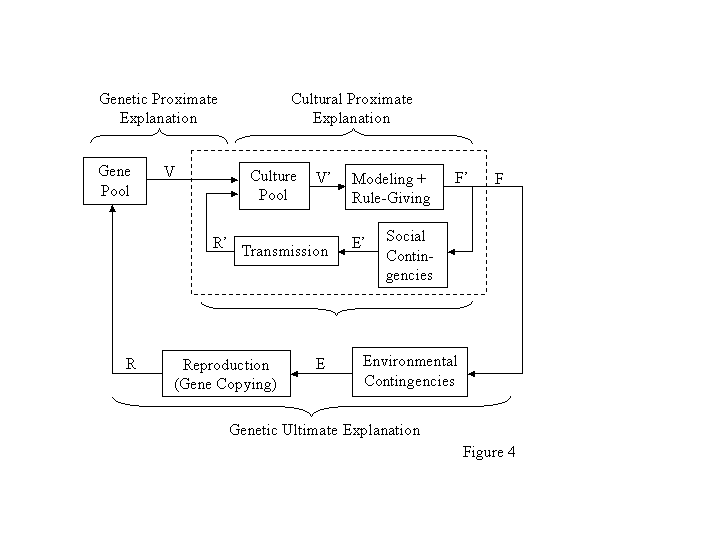
Figure 4 diagrams the relationship between genetic evolution and cultural evolution and the different focuses of their proximate and ultimate explanations. It shows two feedback loops, an outer loop symbolizing genetic evolution and an inner loop (enclosed in the box) symbolizing cultural evolution (cf. Burgos, 1997). The inner loop takes the place of ontogeny in Figure 2B or expression in Figure 2A. The culture pool may be thought of as all the customs present in the groupthe potential variants, resulting in a distribution V. One might be tempted to think of V as a distribution of memes. Our ignorance of the workings of the brain suggests that that temptation is best resisted (Baum, 2000). The mechanism of expression, labeled Modeling + Rule-Giving, embodies the vagaries of the social environment that transform V into a distribution F of actual cultural variants (customs). Models and rules are stimuli that may induce imitation and rule-following, behavior that may be reinforced or punished by group members (Baum, 2000). Alternatively, one could think of the culture pool as directly producing F. (As before, F could also represent the relative frequency of a particular custom.) The structure of the social environment, particularly social contingencies, results in a distribution E of social consequences. For example, food sharing might result in approval, whereas food hoarding might result in disapproval or aggression. From E, transmission results in a distribution R of recurrences that feeds back to change or maintain the composition of the culture pool. The means of transmission are imitation and instruction (Baum, 1994; 2000). They constitute the locus where Boyd and Richersons (1985) indirect and frequency-dependent biases operate.
Figure 4 depicts the way in which cultural evolution and genetic evolution may interact. The distribution F interacts as F in the external loop (genetic evolution) with environmental contingencies to produce distribution Ethat is, cultural practices may affect genetic evolution (Boyd & Richerson, 1985). If, for example, group members who shared food were more likely to marry, then any genes that promoted tendency to share food would be selected.
The braces in Figure 4 indicate the different focuses of proximate and ultimate explanations in genetic and cultural evolution. Genetic proximate explanation focuses on the mechanisms by which genetic variants (distribution V; shown as input to the cultural pooling process) affect cultural evolution. Cultural proximate explanation focuses on the mechanisms (possibly in the nervous system, but specifically related to behavior in groups) by which cultural variants (distribution F) are expressed. Modeling and exhortation, for example, might induce food sharing. Cultural ultimate explanation focuses on the iterative feedback from social consequences that shapes the culture pool over time. Once induced, for example, food sharing might be reinforced by approval, status, or reciprocation. Genetic ultimate explanation focuses on the feedback from environmental consequences of culture on the composition of the gene pool. It represents the feedback of culture on genes that prompts Boyd and Richerson (1985) to speak of gene-culture co-evolution. Failure to keep these four different types of explanation distinct is likely to result in confusion. For example, development (genetic proximate explanation) might account for the presence of imitation and instruction, which underlie cultural ultimate explanation of cultural change over time. Cosmides and Toobys (1992) idea that cultural variation can be explained by genetic expression probably arises from confusing these two types of explanation.
C.7. Proximate and ultimate operant explanations
In operant evolution (i.e., shaping), the distinction between proximate and ultimate explanations is the distinction between physiological mechanism and history of reinforcement. Advantageous behavior is defined by reinforcement and punishment. Explaining the occurrence of advantageous behavior may refer to events in the nervous system or to the history of advantage. As with the other two evolutionary processes, a question like, Why does Liz brush her teeth before she goes to bed? has two interpretations. The proximate interpretation focuses on the mechanism: On any particular night, what causes Lizs tooth-brushing? The proximate explanation would focus on stimuli that regularly precede going to bed and tooth-brushing and events in the nervous system that result from these stimuli and cause tooth-brushing at that time. The ultimate interpretation focuses on a history of selection: How did it come about that Liz brushes her teeth before bed? or What advantage has Liz derived for brushing her teeth? The ultimate explanation focuses on the differential consequences of tooth-brushing in Lizs life that selected tooth-brushing at bedtime over other behavior that might have occurred at bedtime. It would refer to bedtimes in Lizs childhood, her fathers exhortations, reprimands, and approval, and the later incorporation of tooth-brushing into the pattern of behavior surrounding bedtime combined with events at the dentists office. Different stories might be told, but they would all refer to the advantages of tooth-brushing over time.
C.8. Nesting of operant evolution within cultural evolution
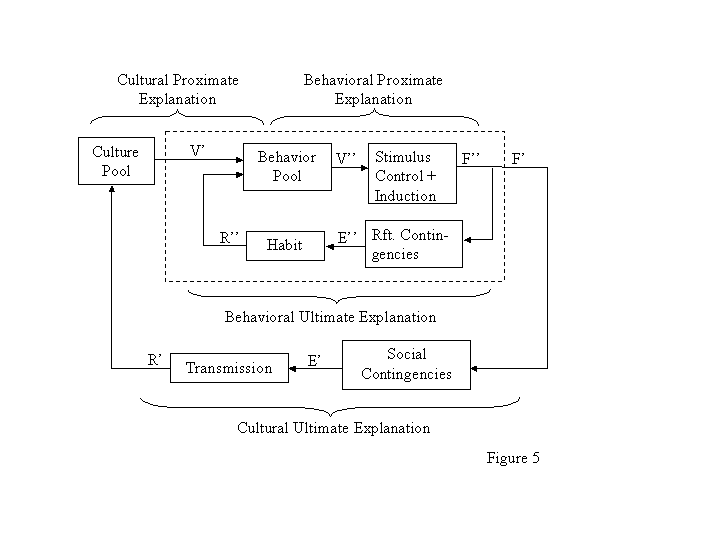
Figure 5 parallels Figure 4, depicting the relationship between operant evolution and cultural evolution. The inner loop, operant evolution, contains the same elements as in Figure 2A. It stands in the place of Modeling + Rule-Giving in Figure 4 or Expression in Figure 2A. The behavior pool consists of all the individuals potential behaviorspecies-specific behavior plus all the behavior ever expressed in this individual. It results in a distribution V of behavioral variants. The culture pool adds to the individuals behavior pool via the distribution V. The mechanisms of stimulus control, including induction, transform V into a distribution (or relative frequency) F of presently occurring behavior. Those mechanisms correspond to what Staddon and Simmelhag (1971) called principles of variation. The structure of the environment, in the form of contingencies of reinforcement and punishment (or reinforcement and punishment feedback functions; Baum, 1973), converts F into a distribution E of environmental effects, or consequences. These reinforcers and punishers constitute the relatively immediate tokens of ultimate reproductive success discussed earlier. Distributed differentially over behavioral variants, they result via mechanisms in the body, particularly in the nervous system, labeled habit in Figure 5, in a distribution R of (relative) recurrences, which in turn changes or maintains the composition of the behavior pool. The behavioral proximate explanation focuses on the stimuli and physiological mechanisms that cause the behavior on particular occasions. The behavioral ultimate explanation refers to the selective effect that the token consequences have on the mechanism and behavior over time. A proximate explanation of sharing of the meat with other group members would point to the occasion of returning to ones village with a captured animal. The ultimate explanation would point to the history of exhortation in favor of sharing, social reinforcement for sharing, and punishment of hoarding. The effect of differential consequences on individuals behavior provides the means for the cultural effects that Boyd and Richerson (1985) call guided variation and direct bias. Guided variation names the contribution of individual innovation to the culture pool. For example, the individual who makes a better arrowhead may catch more food personally, but the method also may be incorporated into the culture pool by imitation and instruction. As we have already seen, direct bias is the effect of the token consequences on transmission of practices. The method of making the better arrowhead personally benefits anyone who uses it. The effect of behavioral evolution on cultural evolution is symbolized by the output of distribution F as F to cultural contingencies, which in turn produce E, the distribution of social consequences.
Taken together, Figures 4 and 5 suggest two levels of nesting. Evolution of individual operant behavior is nested within cultural evolution, which is nested within genetic evolution. To understand human behavior, one needs to attend to all three levels and to all the various proximate and ultimate explanations they imply.
In a non-cultural species, the behavioral inner loop is directly nested within the genetic outer loop (cf. Burgos, 1997). In that situation, gene expression affects the mechanism that causes the behavior and is sensitive to feedback from the token consequences. As in cultural evolution, the token consequences affect gene copying, which feeds back on the composition of the gene pool, favoring alleles that promote the mechanism that allows operant evolution in the first place. If we ask why a laboratory rat presses a lever in an operant chamber, the answer might begin with an explanation of why the rat is so constructed that a contingency between presses and food pellets affects its behaviori.e., the advantages of learning by reinforcement (Baum, 1994; Donahoe, 1999; Zeiler, 1999).
C.9. An example: Self-control
Behavior analysts have progressed on the problem of understanding self-control. Initially, it was analyzed as a conflict between deferred and immediate consequences (Ainslie, 1974). Self-control consists of choosing behavior that pays off in the long run over impulsiveness, behavior that pays off in the short run. For an alcoholic, acceptance of a drink is immediately reinforced, whereas abstinence, though it has a greater reward, is reinforced only at a great delay. The question remains, however, as to how the deferred consequences ever overcome the influence of the relatively immediate ones. How does self-control ever predominate? Rachlin and Green (1972) suggested one possibility: commitment. If one acts at an early time to prevent choosing impulsiveness (e.g., having part of ones salary deposited directly to a savings account before one can spend it), then self-control becomes more likely when the choice arises. Most situations calling for self-control, however, offer no opportunity for commitment, because occasions for impulsiveness are frequent and unpredictable. Some alcoholics take drugs that produce noxious effects when alcohol is in the bloodstream, and some dieters have their jaws wired shut, but most people rely on other means.
Rachlin (1995) argued that self-control might be better understood if it were seen as a pattern of choices extended over time. Occasional impulsiveness (defection from the overall pattern) might occur without necessarily disrupting the extended pattern of doing the right thing. The temporally extended pattern we call eating a healthy diet constitutes self-control even if it is peppered with minor defections like having an ice cream cone. The extended pattern, Rachlin argued, has a higher valuei.e., produces greater reinforcementthan any defection. Those greater and temporally extended consequences explain the persistence of the pattern.
Rachlins (1995) account stops, however, at an awkward place. Although it helps to frame the initial question, it leaves the question unanswered: How does the pattern of self-control ever emerge and persist in the face of frequent opportunities for gaining immediate reinforcement for impulsiveness? In comparison with the contingencies favoring impulsivenesspowerful because of the relative immediacy of the reinforcementthe contingencies favoring self-control would be weak and vulnerable because of their temporal extendedness, as in our comparison to group selection. How could the weak selection ever prevail?
That question raises at least two others: (a) Why would an individual care about eating a healthy diet in the first place? and (b) Why would such a concern spread and persist among many members of a society? On the surface, the answers might seem obvious: quality of life and longevity. Quality of life, however, is a social construct, requiring explanation itself, and death cannot punish nor can longevity reinforce any subsequent operant behavior. Instead, the two questions may be interpreted as being about cultural and genetic evolution: (a) Why do other members of a group promote and enforce eating a healthy diet? and (b) How are genes selected that support acquisition and maintenance of behavior like eating a healthy diet? The answer to the first question requires identifying self-interest on the part of one member in other members health. Such is near at hand, because practices concerning medicine and public health in advanced societies today are costly for the entire group (taxpayers, at least). The contingencies involved may be compared to those involved in so-called altruism. We sacrifice to benefit others if greater benefit accrues to us eventually. The sacrificial behavior is dangerous, however, because of the possibility of cheatingi.e., someones reaping benefits without paying the cost. When most members contribute to the cost of maintaining health in all, poor health is analogous to cheating. Customs promoting and enforcing health-enhancing behavior are selected in such a social setting (see social contingencies in Figures 4 and 5). A possible way that genes might be selected that support eating a healthy diet would be in response to variability in foods available from one environment to another. The advantages to imitation and easy instruction discussed earlier would apply to dietary habits. If you are an immigrant or child newly arrived in a group, the diet eaten by those around you is more likely to promote health than one you choose for yourself. Genes favoring imitation and easy instruction would be selected.
These suggested explanations are incomplete and may be incorrect, but they illustrate the sort of ultimate operant, cultural, and genetic explanations that an understanding of self-control requires. Many of the ideas are open to empirical study. They point to directions in research on behavior within an evolutionary framework.
D. CONCLUSION
Viewed from a sufficiently general perspective, such
as diagrammed in Figure 2A, genetic, cultural, and operant evolution may
all be seen as examples of the same sort of evolutionary process. Since
all three allow the distinction between immediate causation of behavior
and the historical origin of the causes, all three allow both proximate
and ultimate explanations. Indeed,
full understanding requires both proximate and ultimate explanations. The
idea that the three processes are nested in the way suggested in Figures
4 and 5 means that the study of culture needs not only to take account
of cultural evolution, but to be aware of the constraints imposed by genetic
evolution and the contributions of operant behavior and its evolution to
cultural evolution. Likewise,
the study of human behavior needs to take account of the constraints imposed
by both cultural evolution and genetic evolution. A
complete explanation of a behavioral pattern needs to go beyond the processes
that govern individual behavior. It
needs to specify the provenance of behavior in cultural customs and in
genetic effects on development and to explain the origins of effective
stimuli and consequences in cultural and genetic evolution.
ACKNOWLEDGEMENT
I thank M. Ghiselin, S. Glenn, P. Richerson, and H. Rachlin for helpful comments on an earlier draft.
REFERENCES
Ainslie, G. W. (1974). Impulse control in pigeons. Journal of the Experimental Analysis of Behavior, 21, 485-489.
Alcock, J. (1993). Animal behavior (5th ed.). Sunderland, MA: Sinauer Associates.
Alessi, G. (1992). Models of proximate and ultimate causation in psychology. American Psychologist, 47, 1359-1370.
Ashby, W. R. (1956). An introduction to cybernetics. London: Methuen & Co.
Baum, W. M. (1973). The correlation-based law of effect. Journal of the Experimental Analysis of Behavior, 20, 137-153.
Baum, W. M. (1976). Time-based and count-based measurement of preference. Journal of the Experimental Analysis of Behavior, 26, 27-35.
Baum, W. M. (1994). Understanding Behaviorism: Science, Behavior, and Culture. New York: HarperCollins.
Baum, W. M. (1995a). Introduction to molar behavior analysis. Mexican Journal of Behavior Analysis, 21, 7-25.
Baum, W. M. (1995b). Rules, culture, and fitness. The Behavior Analyst, 18, 1-21. (N)
Baum, W. M. (1997). The trouble with time. In L. J. Hayes & P. M. Ghezzi (Eds.), Investigations in behavioral epistemology (pp. 47-59). Reno,NV: Context Press.
Baum, W. M. (2000). Being concrete about culture and cultural evolution. In N. Thompson & F. Tonneau (Eds.), Perspectives in Ethology (Vol. 13, pp. 181-212). New York: Kluwer Academic/Plenum.
Baum, W. M., & Heath, J. L. (1992). Behavioral explanations and intentional explanations in psychology. American Psychologist, 47, 1312-1317.
Baum, W. M., & Rachlin, H. C. (1969). Choice as time allocation. Journal of the Experimental Analysis of Behavior, 12, 861-874.
Benedict, R. (1934). Patterns of culture. New York: New American Library.
Boyd, R., & Richerson, P. J. (1985). Culture and the Evolutionary Process. Chicago: University of Chicago Press.
Boyd, R., & Richerson, P. J. (1992). Punishment allows the evolution of cooperation (or anything else) in sizable groups. Ethology and Sociobiology, 13, 171-195.
Burgos, J. E. (1997). Evolving artificial neural networks in Pavlovian evironments. In J. W. Donahoe & V. Packard Dorsel (Eds.), Neural-networks models of cognition (pp. 58-79). New York: Elsevier Science.
Campbell, D. T. (1965). Variation and selective retention in socio-cultural evolution. In H. R. Barringer, G. I. Blanksten, & R. W. Mack (Eds.), Social change in developing areas: A reinterpretation of evolutionary theory (pp. 19-49). Cambridge, MA: Schenkman.
Cosmides, L., & Tooby, J. (1992). Cognitive adaptations for social exchange. In J. H. Barkow, L. Cosmides, & J. Tooby (Eds.), The adapted mind: Evolutionary psychology and the generation of culture (pp. 163-228). New York: Oxford University Press.
Dawkins, R. (1989a). The extended phenotype: The long reach of the gene. Oxford: Oxford University Press.
Dawkins, R. (1989b). The Selfish Gene (New ed.). Oxford: Oxford University Press.
Diamond, J. M. (1992). Diabetes running wild. Nature, 357, 362-363.
Donahoe, J. W. (1999). Edward L. Thorndike: The Selectionist Connectionist. Journal of the Experimental Analysis of Behavior, 72, 451-454.
Gilbert, R. M. (1970). Psychology and biology. The Canadian Psychologist, 11, 221-238.
Glenn, S. S., Ellis, J., & Greenspoon, J. (1992). On the revolutionary nature of the operant as a unit of behavioral selection. American Psychologist, 47, 1329-1336.
Guerin, B. (1997). How things get done: Socially, non-socially; with words, without words. In L. J. Hayes & P. M. Ghezzi (Eds.), Investigations in behavioral epistemology (pp. 219-235). Reno,NV: Context Press.
Hull, D. L. (1988). Science as a process: An evolutionary account of the social and conceptual development of science. Chicago: University of Chicago Press.
Mayr, E. (1959). Darwin and the evolutionary theory in biology. In B. J. Meggers (Ed.), Evolution and anthropology: A centennial appraisal (pp. 1-10). Washington, D. C.: Anthropological Society of Washington.
Mayr, E. (1961). Cause and effect in biology. Science, 134, 1501-1506.
Mayr, E. (1970). Populations, species, and evolution. Cambridge, MA: Harvard University Press.
Rachlin, H. (1994). Behavior and mind: The roots of modern psychology. New York: Oxford University Press.
Rachlin, H. (1995). Self-control: Beyond commitment. Behavioral and Brain Sciences, 18, 109-159.
Rachlin, H., & Green, L. (1972). Commitment, choice, and self-control. Journal of the Experimental Analysis of Behavior, 17, 15-22.
Richerson, P. J., & Boyd, R. (2000). Built for speed: Pleistocene climate variation and the origin of human culture. In N. Thompson & F. Tonneau (Eds.), Perspectives in Ethology (Vol. 13, pp. 1-45). New York: Kluwer Academic/Plenum.
Ringen, J. (1999). Radical behaviorism: B. F. Skinner's philosophy of science. In W. O'Donohue & R. Kitchener (Eds.), Handbook of Behaviorism (pp. 160-178). New York: Academic Press.
Seeley, T. D. (1989). The honey bee colony as a superorganism. American Scientist, 77, 546-553.
Simon, H. A. (1990). A mechanism for social selection and successful altruism. Science, 250, 1665-1668.
Skinner, B. F. (1938). Behavior of organisms. New York: Appleton-Century-Crofts.
Skinner, B. F. (1953). Science and human behavior. New York: Macmillan.
Skinner, B. F. (1957). Verbal Behavior. New York: Appleton-Century-Crofts.
Skinner, B. F. (1969). The phylogeny and ontogeny of behavior. In Contingencies of reinforcement: A theoretical analysis (pp. 172-217). New York: Appleton-Century-Crofts.
Skinner, B. F. (1981). Selection by consequences. Science, 213, 501-504.
Staddon, J. E. R. (1973). On the notion of cause, with applications to behaviorism. Behaviorism, 1(2), 25-63.
Staddon, J. E. R., & Simmelhag, V. (1971). The "superstition" experiment: A re-examination of its implications for the principles of adaptive behavior. Psychological Review, 78, 3-43.
Tooby, J., & Cosmides, L. (1992). The psychological foundations of culture. In J. H. Barkow, L. Cosmides, & J. Tooby (Eds.), The adapted mind: Evolutionary psychology and the generation of culture (pp. 19-136). Oxford: Oxford University Press.
Wilson, D. S. (1999). Evolutionary psychology: The new science of the mind, by David M. Buss. Evolution and Human Behavior, 20, 279-287.
Zeiler, M. D. (1999). An odyssey through learning and evolution. Mexican Journal of Behavior Analysis, 25, 259-272.
Figure Captions
Fig. 1. Population, frequency, and recurrence. Top: a population with 4 substitutable variants, A, B, C, and D. Middle: Frequencies and recurrences of the variants. Bottom: Frequency and recurrence with continuous variation.
Fig. 2. A: The general evolutionary process as a flow chart. A distribution of substitutable variants, V, is transformed by expression into a distribution of interactors, F, which an external standard transforms into a distribution of effects, E, which recurrence transforms into a distribution of recurrences, R, which enters or replaces the pool to produce a new transform V. B: The general evolutionary process chart applied to genetic evolution. A distribution of alleles (V) is transformed by ontogeny (e.g., culture) into a distribution of phenotypes (F), which environmental contingencies transform into a distribution of environmental effects (E), which gene copying transforms into a distribution of recurrences (R), which up-dates the gene pool and produces a new distribution of alleles.
Fig. 3. Illustration of the transformations of Figure 2. In generation i, distribution V contains 3 substitutable variants, A, B, and C, with equal frequencies. The input N has 6 levels, N1 N6, which enter the transformation to F, a distribution of interactors that contains 4 different interactors, a, b, g, and d, in unequal frequencies. This is transformed to a distribution of effects, labeled W, X, Y, and Z to correspond with a, b, g, and d. In the example, the effects for A, W1 W4, sum to 10, the effects for B, W5 X3, sum to 6, and the effects for C, X4 Z1, sum to 8. These sums determine the frequencies in the distribution of recurrences R. A new variant (accident), D, also arises in R. In this example, the recurrence distribution for generation i becomes the distribution of substitutable variants for generation i+1.
Fig. 4. The cultural evolutionary process as an example of the general process (Figure 2A) and as nested within the genetic evolutionary process. The rectangle of broken lines demarcates the cultural process. A distribution of all cultural variants (practices) available V is transformed into a distribution of actually occurring variants F by induction (e.g., modeling) and rule-giving (instruction). The distribution F comes into contact with social contingencies, which transform it into a distribution of social effects (reinforcement and punishment), E. The distribution of effects is transformed by transmission into a distribution of recurrences of the practices R. The distribution of genetic variants that interact with culture V enters the culture pool as input, thus affecting V. The distribution of actually occurring practices F corresponds also to the distribution F that is the operand for environmental contingencies that form the context for the cultural process. The braces indicate the focuses of the different proximate and ultimate explanations.
Fig. 5. The operant (behavioral) evolutionary process
as an example of the general process (Figure 2A) and as nested within the
cultural evolutionary process. The
rectangle of broken lines demarcates the operant process. A
distribution of all behavioral variants available V is transformed
into a distribution of actually occurring variants F by stimulus
control and induction (e.g., by releasers). The
distribution F comes into contact with reinforcement contingencies,
which transform it into a distribution of effects (reinforcement and punishment),
E. The distribution
of effects is transformed by habit (i.e., physiological mechanisms) into
a distribution of recurrences of the behavioral variants R. The
distribution of cultural variants that interact with individual behavior
V enters the behavior pool as input, thus affecting V. The
distribution of actually occurring behavior F corresponds also
to the distribution F that is the operand for social contingencies
that form the context for the individual process. The
braces indicate the focuses of the different proximate and ultimate explanations.