
GENERATION OF CONCEPTUAL
GISTS OF ABSTRACT PATTERNS
John Beeckmans
The University of Western Ontario, London, CANADA, N6A 5B9
Email: jbeeckmans@eng.uwo.ca
KEYWORDS: perception, vision, gist, information, conceptual short-term memory, image.
ABSTRACT
A short study was made on the generation of conceptual gists of two abstract stimuli containing either 66 or 200 circles in a range of sizes arranged in symmetric patterns. Distributions of estimates the numerousness of the circles, made by groups of subjects following either 95 ms or 2 s exposures had realistic average values, and were comparable between groups, indicating that gists of the principal geometrical features of the stimuli were captured and consolidated into long-term memory on the basis of a single fixation. Implications of this finding for the controversy on the nature of the coding of images and for the minimalist vision hypothesis are discussed.
INTRODUCTION
Gists of visual information are continually generated by rapid unconscious processes as we survey our environment. A gist in this sense is defined as a conceptual summary of the principal semantic features of the perceived scene. The most definitive evidence for the existence of gists comes from rapid serial visual presentation experiments in which subjects are first either shown a target picture of a scene, or given a verbal description. Subsequently they are exposed to a rapidly-presented series of pictures (at rates of up to 8 per second), which may or may not contain the target image, and instructed to press a switch when they recognize the target. Several such studies have shown that subjects can recognize target images under these conditions with reasonable accuracy (Potter, 1975, 1976; Intraub, 1981). The fact that fleetingly-glimpsed target pictures can be recognized on the basis of only a verbal description (e.g. "a road with cars") suggests that gists are more conceptual than pictorial in nature. This was confirmed by the fact that target pictures could be detected even with category cues (e.g. an animal) or negative cues (e.g. "a picture that is not of house furnishings" placed within a sequence containing household furnishings) (Intraub, 1981).
These experiments also showed that gists of natural scenes are generated in about one tenth of a second. However, poor subsequent recognition memory for distractor scenes indicates that it takes longer than that to consolidate gists into memory. It is worth noting that the inability to subsequently recognize distractor scenes is not due to backward masking by the succeeding image, because single briefly-presented images immediately succeeded by a distractor image that subjects are familiar with and told to ignore are subsequently reliably recognized (Intraub, 1980). Since poor post-hoc recognition memory for distractor images following rapid serial presentation is due neither to lack of time for gist formation nor to backward masking it must be due time constraints on consolidation into memory.
Although conceptual gists of scenes can be generated in about 100 ms and consolidated into memory in about 300 ms, consolidation of visuospatial aspects can take considerably longer. Intraub (1980) tested recognition memory of groups of subjects who had viewed scenes for 110 ms, each followed by a blank interstimulus interval which varied from group to group. Subjects' recollections were subsequently tested using either true versions of the experimental scenes or their mirror reversal (none of the test scenes contained alphanumeric characters). At the conclusion of the recognition tests the subjects were informed that some of the images they had identified were mirror-reversed, and were asked to identify them. The proportion of correct mirror-reversals reported was found to depend on the interstimulus interval, up to a value of about a second, demonstrating that consolidation of the visuospatial aspects of the targets into memory could continue for up to a second.
Potter (1993, 1999) has proposed that the fleeting conceptual representations that arise in perceptual processing (i.e. gists) constitute a form of memory buffer that she called conceptual short-term memory (CSTM). CSTM is distinct from normal short-term memory, which has a retention scale of seconds or minutes. It is in fact closely linked to long-term memory (LTM), since the virtually instantaneous conceptual parsing of incoming sensory information into gists requires activation of long-term memories. Information in CSTM generally has a very short duration, with only that part which can be organized immediately into a meaningful structure being retained more than momentarily. Thus, as indicated by the greater accuracy of detection over retention in rapid sequential visual presentation experiments, we must distinguish between gists that are retained in memory and those which exist only fleetingly.
Attention plays an important role in gist formation and consolidation. Firstly, it can be focussed on particular regions of a picture, the content of the resulting gist depending on the degree of focussing and its location, and also to set the level of detail consolidated in a gist (Austen & Enns, 2000) . Secondly, attention can be used as a filter to analyze an image according to predefined criteria (which is how target images are recognized). Finally, attention plays a crucial role in determining whether or not a gist of a briefly presented image becomes consolidated into LTM (Intraub, 1984).
To date studies on gists have focussed on recognition of objects, features, items and/or meaning in natural scenes (Potter, 1975, 1976; Intraub, 1980, 1981); words (Lawrence, 1971); sentences (Forster, 1970; Potter et al, 1986); and alphanumeric characters (Sperling et al, 1971; Chun & Potter, 1995). The gists resulting from such stimuli consist primarily of symbolic representations of the items represented, together with, as appropriate, information on factors such as their relative positions, orientations and sizes, and semantic information pertaining to the meaning of the stimulus as a whole. However, gists of complex abstract figures with repeating features would be expected to be very different from those arising from natural scenes, words or characters. The concepts required to represent such stimuli would concern the geometrical relationships and numerousness of their parts more than to the identity of the items represented. The results of the present study, which used abstract stimuli, bear on two issues in the psychology of vision: (1) the imagery debates, about whether images are stored in depictive or conceptual form (see especially Kosslyn, 1994 and Pylyshyn , 1973; 1981); and (2) the minimalist thesis that relatively little information is gleaned from a single retinal image, and that our impression of seeing a rich and detailed environment is due to a process of cumulative acquisition of cognitive information over numerous saccades (e.g. Churchland et al., 1994; O’Regan, 1992).
EXPERIMENTS WITH ABSTRACT PATTERN STIMULI
The main purpose of the short experimental program described below was to determine the nature, extent and accuracy of subjects’ gists of briefly presented stimuli containing numerous colored circles. The content of the gists was determined by having subjects describe what they remember having seen, and by having them estimate the total number of colored circles. The distribution of these estimates from groups of subjects was subsequently compared with those made by other groups who had viewed the stimuli for 2s.
Stimuli
The experiments involved two stimuli (Figs. 1 & 2) displayed on a computer monitor. They consisted, respectively, of 200 and 66 tightly packed, colored circles arranged in a concentric pattern on a black background. In the central zone the circles were of equal size, and outside this region size increased linearly with eccentricity. The images were designed to be viewed at a distance of 0.5m, giving an angular diameter of 3.4 degrees for the central (foveal) zone and an angular diameter of about 20 degrees for the image as a whole. Under these conditions the sizes of the circles in the central regions in the two figures were, respectively, twelve and twenty times the size of the smallest readable character at the center of the fovea (2.8 minutes of arc).
Anstis (1974) showed that character recognition acuity decreases linearly with eccentricity outside of the foveal region, so in order to ensure that all circles were perceived with roughly similar clarity their sizes increased linearly in the radial direction. The image in Fig. 2 contains seven colors including red, yellow, green, magenta, orange, light blue and creamy white, whereas that in Fig. 1 contains in addition purple and two shades of a deeper blue. Colors were distributed pseudo-randomly, that is to say randomly save that placement of the same color in adjacent circles was avoided.
Figure 1. First experimental stimulus (200 circles).

Figure 2. Second experimental stimulus (66 circles).
Procedure
The nature of the information reported by 40 naive subjects exposed for 95 ms to these stimuli was determined. The subjects were told only that they would be asked to report as much detail as possible concerning two multi-colored images they would see very briefly and to estimate the number of elements present (they were not even told to expect circles). Each stimulus was preceded by a small central light grey fixation spot (1s) and, to eliminate or reduce post-exposure iconic images, were immediately followed by a very bright cream-colored blank screen presented for 3s. The subjects then wrote on a form a full description of what they had seen and their estimate for the number of circles. Half the subjects were shown the images in one order, and half in the reverse order. Subjects were faculty, students and staff at the author’s institution and ranged in age from 18 to 65. For comparison purposes similar experiments were conducted with another group of 40 subjects, using 2 s exposures (these subjects were only required to estimate the number of circles). The two sets were well matched in terms of average age (41.1 and 40.8), gender composition (35% and 43% female), and educational level.
Results
With the 95 ms exposures all but two subjects reported seeing numerous circles arranged in a concentric pattern, whereas two thought they saw a hexagonal or honeycomb structure. All but one reported observing several colors, and most named several colors, although a few reported colors (e.g. grey) that were not present in the images. For each stimulus and each presentation time the pooled data for the two orders of presentation yielded essentially linear plots on logarithmic and cumulative probability axes. It was however found that order of presentation had a significant effect, indicating that subjects were influenced in making their estimate for the second stimulus seen by the value previously reported. For this reason it was decided to remove estimates from stimuli presented second from the data set. In addition, there were considerable inter-subject differences in ability to perceive the structure of the images. Since the objective of the program was not to study statistics pertaining to the perception of briefly-presented stimuli but rather to shed light on the contents of single percepts under optimal conditions, it was considered permissible to remove the data from subjects who had difficulty in perceiving stimuli presented very briefly. Accordingly, estimates were only retained for subjects reporting the gradation in circle sizes. The reduced data sets contained 15 points from 95 ms presentations with the first stimulus (Fig. 1) and 11 points with the second stimulus (Fig. 2). The results with the reduced data sets are shown in Figs. 3 & 4 and Table 1. (Ng is the geometric mean estimated number of circles and sg is the geometric standard deviation); it should be noted that in a lognormal distribution 68.3% of the data are in the range Ng/sg to Ngsg . Bracketed figures refer to the unculled data. Culling had negligible effects with Image 1 but substantially raised Ng for Image 2.) The straight lines in the plots are cumulative lognormal functions, using as parameters the geometric mean diameters and geometric standard deviations calculated directly from the respective data sets.
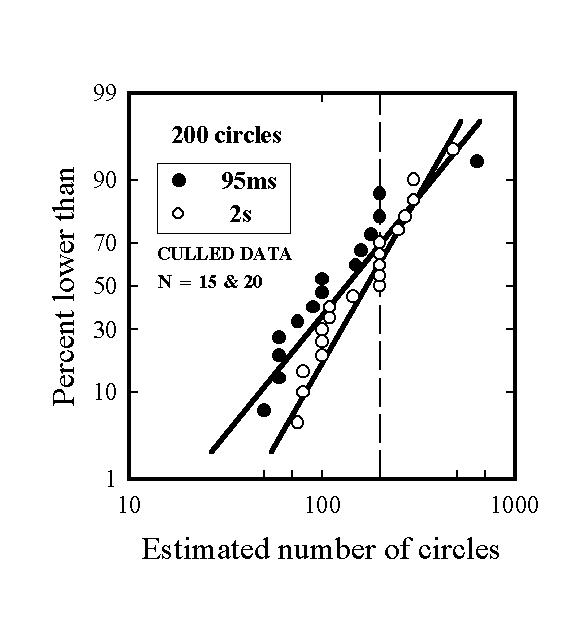
Figure 3. cumulative logarithmic probability plot of estimates of the number of circles in stimulus 1.
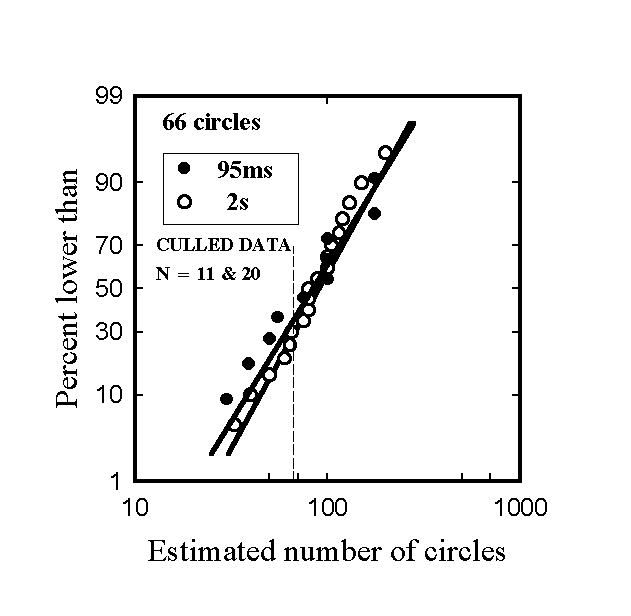
Figure 4. Cumulative logarithmic probability plot of estimates of the number if circles in stimulus 2.
Table 1
Geometric mean parameters for first-seen images
|
95 ms |
2 s |
|
|
Image 1, Ng (N = 200) |
132.0 (133.8) |
169. 8 |
|
sg |
2.23 (2.09) |
1.76 |
|
Image 2, Ng (N = 66) |
84.0 (68.4) |
90.2 |
|
sg |
1.83 (2.06) |
1.72
|
For both stimuli the Ng values with 95 ms presentations were lower, and sg values were higher, than for the corresponding cases with a 2 s exposure, although the differences in means are not significant at the 90% confidence level. Variations in individual estimates may be attributed to two causes: inaccurate perception/memorization of the stimulus, and errors in estimation. If one assumes that the distributions of estimates for large groups of subjects perceiving a stimulus under identical conditions are reproducible, then differences in distributions of estimates between groups exposed under differing conditions must be due to differences in perception/memorization under these differing conditions. In the present case, the slightly larger values of the geometric standard deviation with stimuli seen for 95 ms suggests that they were perceived and/or remembered somewhat less precisely than the corresponding stimuli seen for 2s. Interpretation of the larger values of Ng found with the 2s exposures is more difficult because with the second stimulus the values of Ng are further from the correct values than are the corresponding values with 95 ms exposures. Nevertheless, the written reports from the majority of subjects exposed to the stimuli for 95 ms indicated that they had perceived most of the gross structural features of the stimuli, and furthermore their mean estimates for the number of circles were quite realistic. The relatively large geometric standard deviations are unsurprising because subjects probably used a variety of methods to arrive at their estimates.
Discussion of results
Unlike the tachistoscopic picture stimuli used in previous studies, which contained relatively small numbers of semantically interesting objects (e.g. Potter (1976), Intraub (1980), Thorpe et al. (1996)), the present stimuli contained numerous similar, simple objects. Thus their significance lay not in the nature of the objects represented but in their interrelationships: their number and geometrical arrangement. And they were at the same time both complex and simple: complex because a large amount of information (in the Shannonian sense) is required to specify the exact color, size and location of each circle, yet amenable to simplified conceptual description because of their approximate symmetry.
The estimates of number of circles were generally made several minutes after viewing a stimulus, and hence were based on memory traces in LTM. Such traces, in accordance with the earlier discussion, must have contained conceptual information activated in CSTM during and possibly immediately after exposure to the stimulus. But they would not have contained the bulk of the information, namely the exact color, size and location of every circle. Instead, conceptual information about the structure of the stimuli was activated in CSTM and retained in LTM (see Box 1 below for a listing of this information), enabling the subjects to generate secondary percepts (mental images) of structural analogs of the primary percept, i.e. mental images which conformed to the structure of the stimuli but without detailed chromatic and locational information. And the results reveal that these secondary percepts were sufficiently accurate as to permit realistic estimates to be made, on average, of the number of circles in the stimuli. The written descriptive statements the subjects made after a 95 ms exposure also showed that a majority had perceived both local and global features of the stimuli listed in Box 1. Although the subjects did not explicitly report the quantitative information concerning relative sizes listed in Box 1, this information must have been available in some form to permit them to make their generally accurate estimates of circle numerousness.
Box 1: typical conceptual gist for stimulus 1
Figure contains a large number of tightly packed, brightly colored circles
Size of circles constant in central region but increases toward the periphery
Overall stimulus figure shape circular; black background
Circular central region:
Size of region: (1/7) x size of whole stimulus
Size of circles: (1/5) x diameter of central region
Concentric, circularly symmetric arrangement of circles in peripheral region:
Circle size increases in proportion to distance from the center
Size of largest circles: (1/10) x diameter of whole stimulus
Colors: red, yellow, green, plus several others
It seems probable that the estimations were facilitated by subitizing (Mandler & Shebo, 1982), which allows immediate apprehension of the number present in arrays of five items or fewer. (Note: unlike previous studies involving subitizing, in which subjects made their estimates during or immediately after viewing a stimulus, in the present study the estimates were based on mental images, and without time pressure). Estimates based on subitizing small areas of the mental images could then be extrapolated over larger areas (a process facilitated by the symmetry of the images). This would have been complicated by the variation in the sizes of the circles, which would require several subitizings and extrapolations on different annular regions of constant circle size. Alternatively, some subjects may have based their estimate on the simpler process of subitizing an area containing circles judged to be of average size, followed by extrapolation over the entire image. It should also be pointed out that with the stimuli used in the present study the sector is the only fractional area that can be accurately extrapolated to the full figure.
The experimental results summarized in Table 1 and in Figs. 3 & 4 show that almost as much information about the nature and structure of the stimuli was obtained from a 95 ms viewing period, producing only a single retinal image, as from a 2s viewing period, long enough to permit several saccades and detailed local inspection. A mask was not used because it was thought desirable to maximize the amount of information consolidated into memory. Furthermore, as discussed earlier, rapid serial visual presentation studies have shown that an image’s gist is captured in just over 100 ms, close to the 95 ms exposure time used in this study. Use of a mask in the present experiments would therefore not have materially affected creation of gists of the stimuli.
It may be noted that subjects were able to recall only a few of the colors present in the stimuli. This may be due to the fact that they had been told prior to exposure that they would be asked to estimate the numerousness of the objects or items in the stimuli they were about to be exposed to. Their attention would therefore have been primed toward extracting information on the number of items present rather than on their colors. But the limited capacity of visual working memory was probably also a factor, since Luck & Vogel (1997) have shown that following tachistoscopic presentation it is possible to retain information about only four colors at one time.
The principal conclusion from these experiments is that with the 95 ms presentations an accurate gist of the structure of the stimuli was rapidly captured from a single retinal image, and that this gist contained both local details concerning circle sizes in the various regions of the stimuli, and global properties such as overall circular shape, annular structure, approximate central symmetry, and some of the colors represented.
GENERAL DISCUSSION
The results show that the brain is capable of generating gists of the geometry of complex, abstract stimuli in a very short period of time, probably in less than 300 ms since the iconic image would have been quickly degraded by the succeeding bright blank screen (Sperling, 1960). Consolidation into memory may have taken slightly longer but would not have been impeded by the succeeding meaningless blank screen (Intraub, 1980). It is important to note that prior to exposure the subjects had no inkling of what they were about to see, and that it is safe to assume that they had never before been exposed to comparable stimuli. Of course they had all previously seen circles, but never a single image containing a large collection of differently-sized circles placed in this particular arrangement. The gists were therefore created ad hoc, so to speak, and were presumably not based directly on memories of similar images. In this respect these experiments were unique in that previous experiments on generation of gists involved stimuli containing small collections of previously-seen, categorizable concrete objects.
The results of the experiments will next be discussed in the context of the imagery debates and of the minimalist view of vision.
What can the results of these experiments contribute to the imagery debates?
The imagery debates concern the way mental images are represented in memory. Essentially, the controversy is over whether the representations use a depictive code based on analog properties of the representational medium, or whether they have a conceptual or propositional basis. A thorough review of the matter is beyond the scope of this article, but extended discussion may be found in Kosslyn (1994), who supports the analog hypothesis, and in Pylyshyn (1973; 1981), who advocates abstract symbolic/conceptual modes of representation of images in memory.
The present results appear to support a conceptual representation in memory of the images of the stimuli used in these experiments. Indeed, a conceptual gist whose content is summarized in Box 1 suffices to account for both the subjects’ written qualitative descriptions of the stimuli and for their collectively accurate estimations of the number of circle they contain. On the other hand, information which might be expected to be preserved were the images of these stimuli coded in memory in template-like fashion, such as the exact color and placement of every circle, was not retained. And it seems unlikely that realistic estimates could have been arrived at on the basis of degraded analog representations. Indeed, the estimation results suggest that subjects must have been able to encode the sizes of the smallest circles in the center just as accurately as those of the largest circles at the periphery. Of course, subjects generated and consciously manipulated analog-like mental images of the stimuli to assist in making their reports, but according to the propositional representation view these images would have been generated on the basis of stored conceptual information. Just as, upon request, we can generate a mental image of a camel wearing sunglasses and a long scarf around its neck, even though we have never seen such an apparition. It should also be noted that propositions need not be verbal (Pylyshyn ,1973, p6). For instance, the propositional information in gists of images, especially when only briefly glimpsed as in the rapid serial presentation experiments, is not intrinsically verbalized.
Nevertheless, while it is true that mental images may be constructed on the basis of propositional recipes, so to speak, their instantiation in conscious mental images may involve primitive elements with analog or pseudo-analog properties. Mental images are a part of consciousness, whose nature is still a complete mystery, and one cannot with confidence reach conclusions about its structure from the anatomy of the brain. Still, the fact that the images appearing on the retinas are topographically mapped on the primary visual cortex, V1, and less accurately on several other cortical regions, may play a role in imbuing mental images with analog properties. Although the metric of cortical regions is distorted (as indeed is the metric on the spherically-shaped retinas), non-metric topological properties of retinal images are accurately expressed on V1. Which may be the origin of putative analog properties of directly perceived or memory-based mental images.
Although the mental images used to arrive at estimates of the number of circles in the stimuli in the present experiments were probably generated from coded propositional information, it seems unlikely that the subjects would have performed as efficiently had they been given the information in Box 1 in written form instead of being visually exposed to the stimuli. This suggests that conceptual or propositional information about spatial aspects of visual stimuli is more efficiently captured by automatic, unconscious perceptual mechanisms than from language-based sources.
Implications regarding the minimalist vision hypothesis
Minimalist vision theories posit that our perception of a rich and detailed environment is an illusion, and that in fact relatively little is seen in a single fixation. The illusion is said to arise from a detailed cognitive map of our environment we develop from an accumulation of information over numerous saccades. The minimalist position is defended in (O’Regan, 1992; Churchland et al., 1994; and Ballard, 1996). A variety of arguments are adduced in support of the theory, for instance that we are not confused by the changes in retinal images produced by saccades and that we are unaware of the presence of the blind spot or of retinal non-uniformities. In addition, the phenomenon of change blindness, in which subjects are frequently unable to see a change in an image made between fixations, is believed to support the minimalist view. The changes can be made either during saccades (Grimes, 1996), or by separating the images by means of a brief blank screen (e.g. Rensink et al., 1997).
The present experiments do not undermine the minimalist thesis, but do suggest that the amount of information that can be garnered from a single fixation, while limited, may not be as minimal as the proponents of the theory may believe. In fact it can even be argued that the experiments support the minimalist position, since the gist information summarized in Box 1 represents only a small fraction of the total amount of potential information in the stimulus. Nevertheless, the entire stimulus image must have been seen in some sense (although not necessarily consciously) to enable the subjects to arrive at generally realistic estimates of the number of circles and to produce accurate descriptions of the geometrical features of such complex stimuli. If minimalist vision refers only to conceptual information extracted from visual stimuli and reported post hoc by subjects, such as that illustrated in Box 1, then it makes sense. On the other hand if mimimalist vision purports to include pre-categorical primary percepts from which gists are derived, or to fleeting conceptual gists prior to consolidation of a fraction of their contents into memory, then the theory is on shaky ground. There is simply too much potential for loss of information between the primary percept or the brief, ephemeral gist produced in 100 ms or so, and the final gist consolidated in long-term memory and used by subjects to make their report, to reach any conclusions concerning amount and nature of the information in the original percept and its immediate gist. With the present experiments this means that Box 1 represents only the bare minimum amount of the information accessed in the first 100 ms or so. On the other hand, the full information in the stimulus, i.e. the exact color and placement of every circle, will have reached the primary visual cortex. Although it is probable that only a fraction of this information will have been captured in the primary gist, even prior to consolidation into memory, it is impossible to know how much of it, and in what form, it existed in the brief percepts experienced by the subjects.
CONCLUSIONS
The evidence from this study indicates that the brain has an extraordinary ability to almost instantaneously generate an abstract, conceptual gist that summarizes the structure of a complex and novel two-dimensional geometric figure consisting of large numbers of individual elements. The only previously–seen geometric feature in the present stimuli is the circle, which not only provided the building-blocks but was also implicitly present in the concentric gradation by size of individual circles outside of the central region. The brain probably accomplished this remarkable feat of generalization by classifying features in the stimuli using the pattern-analysis capabilities of its neural networks. It is known that object identification and recognition are performed in the temporal lobe, and this is doubtless also where the rapid, unconscious and automatic geometric-pattern-extraction facilities of the brain are located. The outputs from the temporal lobe, in this case representing coded information about structural features in the stimulus, would subsequently be stored in memory.
Processing of input signals by neural classifiers occurs in parallel, accounting for the extremely rapid generation of multiple-object conceptual gists of semantically important information in perceived stimuli. Additional processing stages would however be required for accessing semantic information about the identified features and objects and their relationships from long-term memory.
The ability of the brain to recognize particular geometric shapes, such as circles, is of course learned. Learning implies conditioning of synaptic weights in certain regions of the temporal lobe so that the appearance of a shape or object in the visual field activates a particular group of output neurons. The results of this study suggest that several geometric features of a complex stimulus can be extracted in this way in parallel by cortical neural networks and used to efficiently store its principal features in memory while discarding a plethora of inessential details.
REFERENCES
Anstis,S.M. (1974). A Chart Demonstrating Variations in Acuity with Retinal Position. Vision Research, 14, 589-592.
Austen,E & Enns,J.T. (2000). Change Detection: Paying Attention to Detail. PSYCHE, 6(11).
Ballard, A. (1996). On the Function of Visual Representation. In K.Akins (Ed.). Perception, New York, NY: Oxford University Press.
Chun,M.M. & Potter,M.C. (1995). A Two-Stage Model for Multiple Target Detection in Rapid Serial Visual Presentation. Journal of Experimental Psychology: Human Perception and Performance, 21, 109-127.
Churchland,P, Ramachandran,V.S., and Sejnowski,T.J., (1994). A Critique of Pure Vision. In C.Koch & J.L.Davis (Eds.). Large-Scale Neuronal Theories of the Brain. Cambridge, MA: MIT Press.
Forster,K.I. (1970). Visual Perception of Rapidly Presented Word Sequences of Varying Complexity. Perception & Psychophysics, 8, 215-221.
Grimes,J. (1996). On the Failure to Detect Changes in Scenes Across Saccades. In K.Akins (Ed.). Perception, (Vancouver Studies in Cognitive Science, 5, 89 – 109). New York: Oxford University Press.
Intraub,H. (1980). Presentation Rate and the Representation of Briefly Glimpsed Pictures in Memory Journal of Experimental Psychology: Human Memory and Learning, 6, 1-11.
Intraub,H. (1981). Rapid Conceptual Identification of Sequentially Presented Pictures. Journal of Experimental Psychology: Human Perception and Performance, 7, 604-610.
Intraub,H. (1984). Conceptual Masking: the Effects of Subsequent Visual Events on Memory for Pictures. Journal of Experimental Psychology: Learning, Memory and Cognition, 10, 115-125.
Kosslyn,S. (1994). Imagery and Brain: the Resolution of the Imagery Debate. Cambridge, MA: MIT Press.
Lawrence,D.H. (1971). Two Studies of Visual Search for Word Targets with Controlled Rates of Presentation. Perception & Psychophysics, 10, 85-89.
Luck,S.J. & Vogel,E.K. (1997). The Capacity of Visual Working Memory for Features and Conjunctions. Nature, 390, 279-281.
Mandler,G & Shebo,B.J. (1982). Subitizing: An Analysis of its Component Processes. Journal of Experimental Psychology: General, 111, 1-22.
O=Regan,J.K., (1992). Solving the AReal@ Mysteries of Visual Perception: The World as an Outside Memory. Canadian Journal of Psychology, 46, 461-488.
Potter,M., (1975). Meaning in Visual Search. Science, 187, 965-966.
Potter,M., (1976). Short-Term Conceptual Memory for Pictures. Journal of Experimental Psychology: Human Learning and Memory=, 2, 509-521.
Potter,M. (1993). Very Short-Term Conceptual Memory. Memory & Cognition, 21, 156-161.
Potter,M (1999). Understanding Sentences and Scenes. In V.Coltheart (Ed.). Fleeting Memories. Cambridge, MA: MIT Press.
Potter,M., Kroll,J.F., Yachzel,B., Carpenter,E., & Sherman,J., (1986). Pictures in Sentences: Understanding Without Words. Journal of Experimental Psychology: General, 115, 281-294.
Pylyshyn,Z. (1973), What the Mind’s Eye Tells the Mind’s Brain: a Critique of Mental Imagery. Psychological Bulletin, 80, 1-24.
Pylyshyn,Z. (1981). The Imagery Debate: Analogue Media Versus Tacit Knowledge. Psychological Review, 88, 16-45.
Rensink,R.A., O=Regan,J.K. & Clark, J.J. (1997). To See or not to See: the Need for Attention to Perceive Changes in Scenes. Psychological Science, 8, 368-373.
Sperling,G. (1960). The Information Available in Brief Visual Presentations. Psychological Monographs: General and Applied, 74(11), 1-29.
Sperling,G., Budiansky,J., Spivak,J.G., & Johnson,M.C. (1971). Extremely Rapid Visual Search: The Maximum Rate of Scanning of Letters for the Presence of a Numeral. Science, 174, 307-311.
Thorpe,S., Fize,D., & Marlot,C., (1996). Speed of Processing in the Human Visual System. Nature, 381, 520 - 522.