HOW TO MODEL CONSCIOUSNESS IN A
MEMORY EVOLUTIVE SYSTEM?
by A.C. Ehresmann and J.-P. Vanbremeersch
Faculté de Mathématiques et d'Informatique,
33 rue Saint-Leu, 80039 Amiens. France
ehres@u-picardie.fr
Preliminary Draft (April 1999)
0. Introduction
One of the main problems faced by the cognitive sciences is to explain what is consciousness and how it may develop in a nervous system. During a long time, this problem seemed outside the scope of science, and reserved to philosophy. However recent advances in neuroscience have opened new perspectives, and it would be impossible to cite all the authors who have recently offered contributions.
Here we propose a mathematical model of some of the processes implicated in consciousness, in agreement with ideas of neuroscientists such as Changeux, Edelman and Crick. It relies on the theory of Memory Evolutive Systems (or MES), which we have developed these last 14 years in a series of papers. We will just recall the results necessary for the sequel, and refer for more details to some of these papers (listed in Section 12 and denoted by EV; cf. also our Internet site: http://perso.wanadoo.fr/vbm-ehr/ ).
0.1. Memory Evolutive Systems
MES represent a mathematical model to study biological, neural or social systems, based on Category Theory, a domain of Mathematics introduced by Eilenberg and MacLane in 1945. The main notions have been progressively introduced and refined to encompass more and more elaborate problems:
• How to model 'open' systems whose components, with various complexity levels, change in time through exchanges with the environment and through internal modifications such as the formation of new complex objects? This question has led us to introduce (in EV 1987) the notion of an Evolutive System (ES); in it the state of the system at a given time, with its components and their interactions, is modeled by a category; a 'complex' object is represented by the colimit (or inductive limit) of a pattern of linked objects standing for its internal organization; and the changes between successive states by 'transition' functors between the corresponding state categories. The ES is hierarchical if its objects are partitioned into complexity levels, so that an object of level n+1 is the colimit of a pattern of linked objects of level n.
In an ES, the 'archetypal' changes (birth/death, scission/collusion) are modeled by a complexification process with respect to a 'strategy' of which the objectives are to add or suppress objects (external exchanges), and to form complex objects (adjunction of a colimit to a pattern) or to decompose complex objects.
• This raises the problem of selection of strategies in an autonomous system. Whence the introduction (in EV 1989) of the notion of a partial internal Center of Regulation, or Coregulator (CR), able to develop a stepwise 'trial-and-error' learning process, thanks to the formation, at each step, of its own internal (more or less deformed) representation of the system, its landscape, on which a strategy is selected.
The case of a neural system shows that its evolution depends on a whole net of competitive CRs. It has led to the definition (in EV 1990) of Memory Evolutive Systems (MES): a MES is an ES with a net of cooperating/competitive CRs, which develop a hierarchical sub-ES, called Memory, allowing to record past experiences in a flexible manner for a better later adaptation; each CR has its differential access to this Memory. The links between objects have a 'strength' and a 'propagation delay' (represented by real numbers).
• In a MES, each CR acts on its landscape according to its own timescale, but the strategies of the different CRs compete for the common resources of the system, and at each time there is a 'play' among these possibly conflicting strategies, with a risk of fractures for some CRs (EV 1991); whence a dialectics between CRs which are heterogeneous by their complexity level and their timescale, generating the complexity of the system. Time has an essential role through the structural temporal constraints imposed on each CR (characterized in EV 1993), and repair processes develop for them being respected anew if they are transgressed. In particular, this leads to an aging theory for an organism based on a cascade of de/re-synchronizations for more and more complex CRs (EV 1993).
• The reductionism problem can be studied in the frame of MES: an object of level n+1 is always the (n-k)-iterated colimit of a ramification based on the level k < n, but is it also 'reducible' to the level k in only one step, i.e., is it a simple colimit of a pattern of level k? A more fine analysis of the form of the links between objects led us (in EV 1996) to a characterization of such reducible objects, and proved that there exist non-reducible objects if the MES satisfies the Multiplicity Principle: there exist non-equivalent patterns which have the same colimit. In this case, a sequence of complexifications cannot be replaced by a unique complexification with respect to a strategy subsuming the intermediate strategies.
• The preceding results applied to a neural system allow to describe an 'algebra of mental objects' (in the sense of Changeux 1983), and they explain the development of a procedural memory, and the formation of higher order cognitive processes. They suggest an approach to the mind/body problem: mental states are not simple physical states of the brain (activation of synchronous assemblies of neurons), but they dynamically supervene on such states through the progressive unfolding of a ramification based on the level of those physical states.
0.2. Contents of the article.
The problem studied in the present article is an attempt to model, in the frame of a MES, the notions of Semantics and Consciousness for a CR. The ideas further develop those proposed in preceding papers (EV 1992 and 1994).
The formation of a semantic memory allows to classify items contained in the Memory in terms of 'concepts'. Such a concept is formed in two steps: first a pragmatically 'acted' classification at the level of specific CRs (such as a CR recognizing colors), then this classification is 'reflected' by a higher CR which stores an invariance class under the form of a (projective) limit, the concept (e.g., the color 'blue').
The introduction of more and more abstract concepts increases internal and external communication, and gives more flexibility to the play among the strategies of the various CRs, allowing for a more efficient comportment. It is essential for the development of higher CRs manifesting some kind of 'consciousness'. In the model we propose, a 'conscious' CR is characterized by the capacity of extending its actual landscape to operate:
• a retrospection process, allowing to retroactively find the causes of a fracture by abduction;
• a prospection process, allowing to program strategies for several steps ahead, taking into account former experiences.
These two processes lead to a better adaptation, with an internalization of the notion of time.
In a first part, we consider a general MES. In a second part, we describe more precisely the MES modeling a neural system and we show how the several processes presented in the first part can be implemented in the brain according to present neurophysiological knowledge. Finally in a third part the general ideas are illustrated by a concrete example in which the various processes are translated into natural language.
Acknowledgments: We are grateful to Profs. Jerry Chandler and Brian Josephson for stimulating discussions which have helped precise our approach to biological and nervous systems.
I. Development of a conscious CR
During the evolution of a Memory Evolutive System (MES), under the effects of the dialectics between heterogeneous CoRegulators (CR), higher order structures may emerge, be they more and more complex objects recording former experiences (situations, applied strategies and their result, possibly concepts), or observers/agents operating on them. It may lead to the formation of internal higher and higher CRs, constituted of emerging objects and links. These CRs, with a longer latency delay (their 'actual present'), play the role of associative CRs regulating lower CRs and they have a strong influence on the dynamics of the system, through their powerful participation to the 'play' among the strategies of the different CRs.
1. Some recalls on CoRegulators (CR)
A CR of a MES is a sub-ES whose objects, its agents; have a particular level of complexity, and which has its own discrete timescale. It develops a stepwise trial-and-error learning process, a step extending between two successive times of its timescale. The step is divided into several more or less overlapping phases: during the first one, its actual present, the landscape is formed; then a strategy is selected on this landscape, realized and its results are evaluated.
1.1. Actual Landscape of a CR (cf. EV 1989 and 1991).
The instantaneous landscape of a CR at a time t gathers the information on the external and internal state of the system upcoming to the agents of the CR at t. Among those, some are very weak or temporary, others remain stable during the actual present of the CR; these later ones form its working memory, called its actual landscape. At each step, an adequate strategy is selected on this landscape, and its commands are sent to the corresponding effectors. A fracture occurs if no adequate strategy is found, or if the selected strategy cannot be realized because of recent modifications not observable in the landscape.
The instantaneous landscape at t is modeled by a category whose objects are the perspectives arriving to the agents at t; a perspective of B is an equivalence class of links (or 'aspects') b from an object B of the system to an agent of the CR, for the relation: two links are equivalent if they are correlated by a zig-zag of links between agents. The links from a perspective of B to a perspective of B' are determined by the links from B to B' in the system which are correlated by these perspectives, so that there is a functor, called 'distortion' from the landscape to the system mapping a perspective of B on B. The CR is identified with a sub-category of the landscape. If a pattern p of perspectives has a colimit (or a limit) c in this landscape, there exists an agent A such that c and all the perspectives in p have at least one of their aspects arriving to A.
The actual landscape at t is a category constructed by binding the sequence of instantaneous landscapes formed during the actual present, say from t to t+µ, but preserving only the perspectives coming from levels near the level of the agents and whose aspects preserve a sufficient strength during the present step. Thus one of its object is determined by the successive states of such a perspective from a date between t and t+µ up to the end of the step.
1.2. Intentional CR (cf. EV 1994 and 1996)
For a lower CR, the selection of a strategy in its actual landscape may be imposed by another CR, or directly ensue from the recognition of the situation via the links of the CR with the Memory; the command then comes from the Memory rather from the CR, except that the CR has information on it.
For the CR to really 'select' a strategy, it must have a sub-system of 'evaluating agents' (at least two, say + and -). Each strategy available to the CR already used in similar situations is linked to one of them through the storage of its results, for instance S is linked to + if S has succeeded, the link being strengthened (i.e., its strength increases) each time the result is repeated. There is also a link in the other way (possibly not inverse) corresponding to the command of the effectors of S if the strategy is 'good', to their inhibition if not (so that the strategy would no more be chosen). The comparison between strategies classified by the same evaluating agent (e.g., between 'good' strategies) can be done automatically, using the strength of the links linking the strategies to the evaluating agents.
An associative CR is said to be D-intentional, or intentional in the sense of Dennett (1990), if it has a sub-system of evaluating agents, so that it acts 'as if' it is able to optimize its choice of strategies in its landscape.
The CR will be said intentional (in the full sense and not only 'as if') when the strategies and their comparison have an internal representation in the landscape.
An intentional CR is mainly formed in a MES in which a semantic memory has developed, and then it may select strategies under the form of a concept.
2. Semantics (cf. EV 1992)
A Semantic Memory may gradually develop in a complex enough MES, through the detection of specific invariances by the CRs, which leads to classify memorized items according to their main attributes, and form new formal units representing their invariance classes.
2.1. Formation of CR-concepts
It is a two steps process:
a) Comparison of two objects with respect to a particular attribute: A lower CR, say E, receives in its landscape only some perspectives of the objects recognized by the system; for instance, in a neural system a 'color' CR retains only the color of an object. Thus it operates a 'pragmatic' classification of the objects depending on the attributes it may recognize: Two objects are 'acted' as equivalent if their different aspects are interconnected by the same pattern P of agents of E; for instance, all blue objects activate the same assembly of neurons of the color CR.
If M is an object in the Memory, the pattern P of agents of E which interconnects all the aspects of M for E is called the E-trace of M, or the E-pattern activated by M. Two objects M and M' are said to be in the same E-class if they have the same E-trace. (More formally, M and M' are in the same E-class if they have the same 'shape' for E, in the sense of the "Shape Theory" of Borsuk, 1975.)
b) Formation of a concept which characterizes a class of objects having a common attribute: The above classification can acquire a 'meaning' only at the level of a higher CR, with a longer period, which receives in its landscape information both on the pattern P and on the objects M by which it is activated. This CR can observe the correspondence between P and the E-class of M, and store it under the form of an object C in the Memory, called the E-concept of M, which singles out the E-class of P, i.e., the invariance class of M with respect to the attributes classified by E; for instance, the color-concept 'blue' classifies all the representations of blue objects with respect to their color.
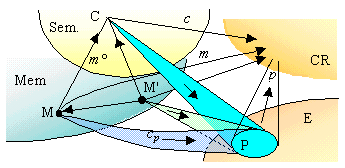
Let us assume there is a CR whose actual landscape contains simultaneously perspectives m of the various objects M of an E-class, a pattern p of perspectives of their E-trace P, and common messages cp from m to p formed by the aspects under which M is seen from E. Then the CR memorizes the E-class of M as a perspective c limit of p in its landscape. The object C source of c is a limit of P in the system, called the E-concept of M. It exists a link m° from M to C binding cp (by definition of the limit of a pattern). Thus C becomes a final object in the E-class of M, and its classification domain represents the E-invariance class of M.
2.2. Semantic Memory.
An object M may have several invariance classes with respect to different CRs, each corresponding to a classification with respect to different attributes. For example a blue triangle and a blue circle have the same color-concept 'blue'; but the shape-concept 'triangle' of the blue triangle represents the class of triangles, while that of the blue circle represents the class of circles. Concepts with respect to several attributes can then be stored as limits of a pattern formed by these different attributes ('blue triangle' is the product of 'triangle' and 'blue'). Then more and more complex concepts are constructed based on these 'concrete' concepts, to form the Semantic Memory. Such a concept can be thought of as an abstract prototype for a class of objects, its various instances, which have a 'family resemblance' (in the sense of Wittgenstein 1953); remark that it does not necessitate the existence of a language.
The Semantic Memory is a sub-ES of Memory whose objects, called concepts, are iterated limits of patterns of CR-concepts. It is obtained through successive complexifications applied to the sub-category of Memory formed by the CR-concepts associated to the different CRs, and their compatible links.
The development of such a semantics in the MES gives more flexibility to the choice of strategies and to the play among the strategies of the various CRs. Indeed, the system will recognize constancies through changing circumstances and react in a specific manner not to a particular situation but to all similar situations; for instance an object is identified whatever be its size or location in the visual field. And the Procedural Memory (formed by stored strategies) will also be classified into concepts of strategies, allowing higher level CRs to select their strategies under the form of such a concept, instead of choosing a specific object of its invariance class. Thus a new degree of freedom is introduced in the formation of the strategy effectively realized on the system, since the play among the strategies may single out the most adapted instance of the concept in view of the strategies relayed by the other CRs. For example the command to hold an object will not activate the same muscles depending on the location of the object, its size, its shape.
3. Conscious CR (cf. EV 1992 and 1994)
For Nietzsche, to become conscious of something is to uncover motivations for it. It is what will do an intentional CR capable of consciousness, called briefly a conscious CR. Such a CR will develop a process which allows an internal trespassing of temporal constraints thanks to a dynamic control of the Memory, leading to more adapted responses; this agrees with ideas of Changeux (1983) and Edelman (1989) on consciousness.
Conscious CRs are integrated in a sub-MES with its own temporality and a differential access to the semantic memory, but with no direct relations with the environment. So they act in a somewhat 'schizophrenic' manner, the relations with the exterior being under the form of imposed fractures.
More precisely, we say that a CR is conscious if it is capable of the 3 following operation which are mutually intermingled (the neurophysiological bases of these operations in a neural system are studied in Part II):
3.1. Retroactive extension of the working memory of the CR
A diffuse activation of the whole system ('awareness'), characterized by an increase of the strength of the links, allows the CR to be informed of some aspects initially not retained in its actual landscape (used as its working memory). This activation can result from a strategy chosen by the CR in view of a finer analysis of the present situation, or be automatically effected because of a fracture.
Then the actual landscape of the CR can be retroactively extended thanks to the recovery of information contained in the instantaneous landscapes from which it is constructed, but not initially retained. Such information may have been bypassed in the actual landscape because it concerns objects with stability spans shorter than the actual present, or depends on the decomposition of complex objects which had been globally identified.
Formally, the extension of the actual landscape at t consists in the construction of a sequence of larger categories, recovering some of the perspectives figuring in the instantaneous landscapes at the different dates of the actual present of the CR at t but not observable in the actual landscape, in particular coming from the Memory. It relies on the two processes described hereafter: retrospection and complex planning.
Thus the extended landscape is a mirror not of the present state but of the various evanescent traces accumulated in the working memory during the actual present.
3.2. Retrospection (abduction to detect the causes of a situation, e.g. of a fracture)
The CR will analyze the extended landscape to find the date of the fracture and/or the probable causes of the situation, taking into account information on similar former situations stored in the Memory. It executes a sequence of loops between the CR and the Memory, in particular the semantic memory, to recognize concepts and/or their instances which have been perceived in the instantaneous landscapes; these are gradually made more precise thanks to recovered attributes, and from them, by an abduction process, the CR retrieves the possible causes of their activation by the situation (e.g., the fracture).
For instance, the sentence "the fish attacked the man" creates a fracture in the (usual) conscious CR; the abduction process finds that the fracture comes from the fact that a 'typical' fish is not aggressive for man; a new abduction evokes dangerous animals related to fishes, whence the recall of a stark, which makes the sentence meaningful. Another example is the presentation of two different objects to the two eyes; the receptors activated by the two objects will activate corresponding concepts, say A and B, in the Semantic Memory, and both can be recovered in the extended landscape; as both have the same strength, they'll be seen alternatively.
The retrospection process is modeled by the stepwise research of final objects in the sub-categories of the successive extended landscapes formed by the perspectives coming from the Memory and representing similar situations or their attributes.
3.3. Complex planning
It divides into two kinds.
a) Prospection process: it allows the selection and longer term planning of strategies possibly covering several steps of the CR. The CR forms successive extensions of its actual landscape to find simultaneously the causes of the situation (cf. above) and the available strategies to respond; and it virtually anticipates (without cost) their possible effects on the future evolution, taking into account information stored in the Memory, including the risks of fractures. This process may lead to a new extension of the landscape (via a new loop between the CR and the Memory) to find more accurate information helping to chose a better adapted strategy.
The prospection process consists in the internal formation, on successive extensions of the landscape, of a sequence of 'virtual' anticipated landscapes on which strategies are 'tested'. It permits to select a sequence of strategies Si for several steps ahead; each strategy depends on the realization of the preceding ones, and the strategy Si+1 will be realized only once Si has been realized. Their successive commands of effectors may be set forth either directly on the landscape of the CR, or indirectly by imposing strategies on lower CRs.
For instance, it will be possible to plan the dynamic formation of a 2-iterated colimit N in two steps: the first strategy will add the colimits Ni of some patterns, thus also introducing simple or complex links between these Ni; then a new step of the CR begins, and its new strategy will add the colimit N of a pattern formed by these Ni and some distinguished links between them.
b) Multiple planning: In a conscious CR, the continuity of a process can be preserved in spite of temporary interruptions followed by resuming the process where it had been stopped (in object-oriented programming languages, this is simulated by a 'thread'; cf. Niemeyer and Peck 1997; also used by Josephson 1998). In some cases, this can be modeled as above by a choice of strategies on several steps ahead, for instance if the interruptions consist in waiting that some operations imposed by one of the strategies be realized by a lower CR.
In other cases, we'll have to simultaneously plan several long term strategies preserving a continuous command of lower level effectors, but perceived at the conscious level only alternatively. For example it is possible to talk while driving a car, each of these activities asking for more or less attention at a given time. The conscious reactivating of one of these strategies, say S, at a given date can be planned by the conscious CR, or imposed to it by a fracture caused by an external stimulus to a lower CR participating in the realization of S (a road obstacle requires the attention of the driver to slow-down). The reactivation of S is instantaneous and does not cause a fracture in the conscious landscape (the discussion is resumed at the same point).
In the extended conscious landscape, the different strategies are modeled by the objects of the multicolimit (in the sense of Diers 1971) of a pattern subsuming the objectives of all the strategies; the specific object which is activated (thus determining the strategy which is realized) at a given time depends on the context.
II. Application to a neural system
(cf. EV 1990)
4. The MES of neurons
A neural system can be modeled by a MES, and, for a higher level animal, this MES will have some conscious CRs in the preceding sense. Let us describe how the different processes involved in the functioning of a MES can be implemented in the brain, taking into account known neurophysiological results. In particular, we'll use the rule of Hebb and the existence of anatomical loops between different brain areas (limbo-cortical and cortico-cortical loops), such as the 'loop of consciousness', on which Edelman (1989, page 100) bases his theory of primary consciousness.
4.1. The category of neurons
To construct the category of neurons at a given time t, we first consider the oriented (multi)graph whose vertices are the neurons and the edges the synapses between them. This graph is labelled as follows:
• To each neuron N we associate its activity at t, measured by its instantaneous spike frequency, and its threshold potential; we say that N is activated at t if its activity increases at t.
• To each synapse from N to N' we associate its strength at t which depends on the propagation delay and on the activity of N needed to activate N' along the synapse.
• The strength of a synaptic path is the product of the strengths of its successive links.
The category of neurons at t has for objects the neurons and for links the equivalence classes of synaptic paths for the relation: two paths between the same neurons are equivalent if they have the same strength. The composition of links is their concatenation. This category is labeled by the strength of the links.
4.2. Synchronous assemblies of neurons
Medical imaging shows that brain functioning relies on the activation of synchronous assemblies of neurons (in the sense of Hebb 1949, or neuronal group for Edelman 1989, cf. also Changeux 1983). Such assemblies will be modeled by 'formal' units added to the category of neurons, and they'll emerge to become the colimit of the pattern of neurons constituting the assembly.
The MES of neurons is a MES in which the state category at t is a category deduced from the category of neurons by a sequence of complexifications. Thus its objects, called cat-neurons, are constructed as iterated colimits, based on the category of neurons.
The cat-neurons of order 1 are obtained as the (simple) colimit of a pattern of neurons, and they model Hebb synchronous assemblies of neurons. Indeed, the emergence by complexification of a colimit N added to bind together a pattern P is manifested by the strengthening of the distinguished links of the pattern, so that the activities of its objects become synchronized, allowing for collective actions. The synchronization lasts only a short time (cf. Rodriguez et al. 1999; Miltner et al. 1999; Usher & Donnelly 1998); it is relative to the complexity of the neurons (e.g., longer in associative cortex) and does not imply that the propagation delays of the distinguished links of the pattern are zero, but only that they are short compared to the latency delays of the objects on which they act.
4.3. Multiplicity Principle
The links from the cat-neuron N, colimit of P, to a (cat-)neuron N' correspond (definition of a colimit) to the collective links from P to N', along which N' can be activated by the synchronous assembly operating as a whole. Thus the cat-neuron N models the synchronous assembly in its capacity to transmit its activation.
However, experimental studies prove that two different synchronous assemblies may transmit the same activation, thus realizing the same 'function'. This property is important, because it is at the root of the robustness of the brain functioning, for instance allowing for recuperation after partial destruction of the brain. To model this property,
We assume that the MES of neurons satisfies the Multiplicity Principle: Two non-equivalent patterns P and P' may have the same colimit; the passage between P and P' is then called a complex switch.
5. Activity of a cat-neuron
5.1. Higher order cat-neurons
Cat-neurons of higher order are obtained by successive complexifications. In particular a cat-neuron M of order 2 is the colimit of a pattern of cat-neurons Ni of order 1; its emergence consists in their synchronization through the strengthening of their distinguished links. As each Ni represents a synchronous assembly of neurons, M represents a synchronous superassembly (i.e., assembly of assemblies) of neurons. Though M is a formal unit, its activation consists in a dynamic physical process, namely the unfolding of this superassembly: first activation of the several synchronous assemblies Ni , and then synchronization of their activities along their distinguished links. For instance this process has been observed in odor encoding (Wehr & Laurent 1996).
More generally, a cat-neuron of order k+1 is the colimit of a pattern P of cat-neurons of order k, acting as a synchronous assembly of cat-neurons. It follows that it is also a k-iterated colimit of a ramification based on the neurons, and thus it represents a synchronous hyperassembly of neurons, i.e., an assembly of assemblies of assemblies of.... of neurons. Its activation consists in the dynamic gradual unfolding of the ramification formed by these successive assemblies from the level of the neurons up.
5.2. Links between cat-neurons
There are two kinds of links between cat-neurons. A 'simple' link between two cat-neurons N and M binds a cluster of links between the cat-neurons of the synchronous assemblies of cat-neurons they represent; its strength is deduced from that of the links of the cluster.
Let P and Q be two patterns representing synchronous assemblies of cat-neurons, N and M the cat-neurons which are their colimits. A cluster from P to Q is a maximal family of links from the objects Pi of P to the objects Qk of Q, correlated by the links of P and satisfying the condition: each Pi is linked to at least one Qk and, if it is linked to several ones, those are connected by a zig-zag of distinguished links of Q.
A (P,Q)-simple link from N to M is obtained as the binding together of the links of a cluster, and its strength is computed as the weighted sum of the strengths of the links of the cluster.
We have said that two different synchronous assemblies P and P' may give rise to the same cat-neuron N (Multiplicity Principe). It follows that a composite of simple links may not be simple; it is then called a 'complex' link.
A link which is not simple is called a complex link if it is the composite of a sequence of simple links binding non-adjacent clusters, separated by complex switches. Its strength is the product of the strengths of these simple links.
5.3. Activity of a cat-neuron
The activity of the cat-neuron of order 1 at a given time depends on the activity of the various neurons of the synchronous assembly of neurons which it binds, and on the strength of the distinguished links along which they exchange information. A higher order cat-neuron N is the colimit of a pattern P of cat-neurons of a lower order. Thus, by induction, we may compute the activity of N assuming that the activities have already been defined at the level of P:
Let (t,t+µ) be the mean propagation delay at the level of the cat-neuron N colimit of a pattern P. For each time y between t and t+µ, an object Pi du pattern P is also activated by the other objects Pj to which it is linked. The activity transmitted to Pi by Pj via a distinguished link is equal to sji(y)cj(y) if cj(y) is the activity of Pj and sij(y) the strength of the link. Summing up, the activity of Pi at y is the sum ai(y) of the activity ci(y) of Pi with the several sji(y)cj(y) corresponding to the distinguished links arriving to Pi. Then the activity a(t) of N will be the means of the sum of the ai(y):
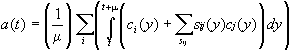
The activity of N will increase if the activity of the cat-neurons Pi increases as well as the strength of their distinguished links. However N can also represent another assembly P' (Multiplicity Principle). In some cases, P might be activated while P' is externally inhibited; then there will be a conflict for the activation de N, the most important effect taking the advantage after some latency time. In particular if the switch between P and P' is complex, the activity of P must be large enough for it to be transmitted to P' via N; in other terms, a complex switch requires some energy to be effected.
6. Extended Hebb rule
6.1. Hebb rule
In 1949 Hebb has proposed that the strength of a synapse is modified in agreement with the following rule (in fact it had already been suggested by Hering in the nineteenth century):
Hebb rule: If the activities of the pre-synaptic and of the post-synaptic neurons are simultaneously increasing, then the strength of the synapse joining them increases at the same rate; conversely, if they vary in opposite ways, the strength of the synapse decreases.
This rule has been experimentally confirmed for synapses in many areas of the brain (e.g., Zhang et al. 1998; Frey & Morris 1997), and also for groups of synapses (Engert & Bonhoeffer 1997). We are going to prove how it generally extends to cat-neurons and simple or complex links between them.
6.2. Hebb rule extended to simple links between cat-neurons
Hebb rule for simple links: The strength of a simple link between two cat-neurons N and M increases if the activities of N and M are simultaneously increasing; it decreases if they vary in opposite ways.
Let us prove this rule for order 1 cat-neurons; it extends by induction to higher order cat-neurons. Indeed, let g be a simple link from N to M binding a cluster G from P to Q. Let us suppose that the activities of N and M are simultaneously increasing by reflection of those of the synchronous assemblies P and Q. The (usual) Hebb rule implies that the strength of a link of the cluster will increase, since the activities of the neurons Pi and Qk which it links increase. The strength of the link g being the weighted sum of the strengths of the links of the cluster, it follows that it also increases. Thus the Hebb rule is extended to simple links.
6.3. Hebb rule extended to complex links
Now let us consider a complex link gh, composite of two simple links g from N to M and h from M to N°, these links binding non-adjacent clusters from P to Q and from Q' to P°, with a complex switch between Q and Q'. Its strength is an increasing function of the strengths of the two simple links. If the activities of N and of N° increase, it seems no conclusion is possible, because we don't know how the activity of M varies. However, if the activity is N becomes large enough to be transmitted to M via g, it follows from the Hebb rule for simple links that the strength of this link will also increase. But then the activities of M and N° simultaneously increase, and the same rule implies that the link h from M to N° is also strengthened. In this case, the strength of the complex link gh will increase. Thus:
The Hebb rule also extends to complex links in the case the activity of the cat-neurons it connects is greater than some threshold.
7. Development of the Memory
An animal has an innate knowledge, or later learns, the main features of its environment and different strategies to cope with it. Its experiences (perceptions, motions, choice of strategies and their results, cognitive processes of any kind) are memorized in a flexible way under the form of neurons (for simple stimuli), or cat-neurons representing synchronous assemblies of neurons, or more complex higher order cat-neurons. They will be recognized in similar situations later on via the activation of these cat-neurons, thus leading to more adapted comportments.
The emergence of such cat-neurons develops the hierarchical sub-ES of the MES of neurons, denoted by Mem, consisting in its Memory. Mem is not an immovable record, but is progressively modified to take into account new experiences; for instance if a once successful strategy later fails because of a change in the environment, this will be reflected in Mem by a weakening of the strength of the link along which this strategy is selected.
7.1. Empirical Memory and Procedural Memory
A simple stimulus activates a pattern R of receptors in lower areas of the brain (e.g., the visual area V1); following Hebb rule, its distinguished links are strengthened through the synchronous activation of the objects of the pattern. If the activation is great enough or if the stimulus is repeated, the pattern is transformed into a synchronous assembly of neurons, which takes its own identity, and is modeled by a cat-neuron M in Mem (emerging as the colimit of R).
A more complex, external or internal, stimulus is decomposed into a pattern of simpler stimuli already recorded under the form of (cat-)neurons Mi , and it is memorized by the emergence of a higher order cat-neuron M in Mem binding the pattern of these Mi.
The memorization of a stimulus Q is modeled by the formation of a cat-neuron M in Mem, representing the synchronous (hyper)assembly of neurons activated by Q. We call M the record of Q.
If a similar stimulus is presented to the animal later on, M will be retrieved from Mem through its activation, and that allows the recognition of the stimulus through the different CRs (cf. Section 8).
In the same way, a strategy will be recorded by the formation of a cat-neuron, via the strengthening of the links of the pattern connecting its objectives. The cat-neurons (of any orders) representing strategies, and the links compatible between them (e.g., those corresponding to the insertion of a sub-strategy into a strategy) form a sub-ES of Mem, called the Procedural Memory and denoted by Strat.
7.2. Semantic Memory
It is formed (as described in Part I) in neural systems with a developed associative cortex, and allows to classify items recorded in Mem into invariance classes with respect to different attributes. Its objects, called concepts, are formal objects C representing the class of (cat-)neurons M which activate a same synchronous assembly of neurons (or, in higher levels, of cat-neurons); we say that such a C is a concept of M (for some attributes), and that M is an instance of C.
A concept is modeled by the limit C of a synchronous assembly of (cat-)neurons, P; its instances are the cat-neurons M in Mem for which there exist a common message from M to the pattern P (able to transmit the activation of M to the whole assembly). This common message is binded into a link from M to C.
The activation of an object M of Mem is transmitted to its concept C, if it exists, and via C to the concepts relative to more particular attributes (color-concept, shape-concept,...). Conversely, the activation of C is transmitted to the synchronous assembly P which it subsumes but not to the several instances of C; however, if there is a diffuse activation of Mem (e.g., following an increasing 'awareness'), a particular instance M might be activated, in which case the link from M to C is strengthened by Hebb rule, allowing an abduction process to proceed from the concept to this instance.
8. Functioning of a CR
The CoRegulators (CR) of the MES of neurons are sub-ES contained in, or based on, various cortical areas (based meaning that their agents are cat-neurons representing synchronous (hyper)assemblies of neurons of these areas); for instance, there are lower CRs contained in the visual area V1, while the agents of higher CRs are more or less complex cat-neurons based on the associative cortex. (Cf. Section 9 for a comparison with the units of treatment and the conscious units of Crick 1994.)
Each CR has a differential access to Mem through its landscape thanks to which it may recognize recorded items. It also participates to the development of this memory, directly and/or indirectly via the play among the strategies of the different CRs. Let us describe how these operations rely on the activation of thalamo-cortico-cortical loops through Hebb rule.
8.1. Storage and retrieval by a CR
A particular CR, say E, may only recognize some attributes of a stimulus. Let us consider a stimulus (external or internal) Q which activates a pattern R of (cat-)neurons. The stimulus will be observable in the landscape of E only through the pattern r' of perspectives coming from a sub-pattern R' (possibly void) of R. Its attributes identified by E will be memorized through the synchronization of r' which strengthens R' into a synchronous assembly of (cat-)neurons. This assembly is stored in Mem under the form of a cat-neuron M', called the E-record of Q.
The stimulus Q has a E-record M' if M' is the colimit of the pattern R' and if there exists a perspective m' of M' which is the colimit of the pattern r' of perspectives activated by Q in the landscape. In this case, the stimulus can be later recognized by E through m', when m' is activated enough to become observable in the actual landscape.
If Q has a E-record M', its later recognition by E is a consequence of Hebb rule. Indeed, since M' has a perspective m' colimit of r', there exists an agent A of E such that both m' and each perspective in r' have some aspects arriving to A. The later presentation of the stimulus will activate A via the aspects of r' and M' via the pattern R'. The activities of A and of M' being simultaneously increasing, Hebb rule implies that the strength of a link from M' to A increases; thus the perspective m' is strengthened, and becomes visible in the actual landscape of E, thus allowing for the recognition of the attributes of Q detected by this CR.
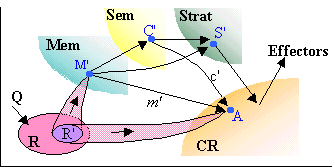
A CR will entirely recognize the stimulus and not only some of its attributes if Q has a CR-record M' identical to the record M of Q, which means that each receptor activated by Q be observable in the landscape of the CR. However we should be cautious that, even in this case, the CR accesses not to the record M itself, but only to its perspective m' in the actual landscape, i.e., to a 'subjective' representation of M (in the case of a conscious CR, could this difference be at the root of the 'qualia'?).
The difference between the record M and its perspective m' explains that it can be difficult to voluntarily retrieve M from Mem at a given time, if m' is then not strong enough to be observable in the landscape; for instance, when we meet somebody we may not find immediately his name though we recall it later on. More severely, it accounts for some memory dysfunctions which come not from a destruction of records, but from the severing of their links to the conscious CR.
8.2. Role of the CRs in the development of Mem
A CR will participate in the memorization of a stimulus Q, directly by its choice of strategies, and/or indirectly through the play among the strategies of the different CRs. Indeed, a higher CR can take as one of the objectives of its strategy to strengthen the pattern r' of perspectives it has received from Q (cf. above); for instance we can make an effort to memorize a phone number. In this way it may promote the formation of the corresponding CR-record M', and eventually also memorize a strategy S' in response to Q, so that the later activation of M' will activate S' via the link from M' to S'.
In many cases, the whole record M of Q will only emerge from the play among the strategies of the different CRs: this play will bind the various CR-records of Q, that leads to the strengthening of the whole pattern R into a synchronous assembly, represented by M. Now if M is linked to a recorded strategy S, this strategy will be activated by Q through M, and the choice of S by the system may impose a strategy to a CR having no CR-record, possibly causing a fracture in its landscape.
Higher CRs also participate in the development of the semantic memory, and can effect the retrieval of a stimulus Q through a concept C of M (cf. Part I).
8.3. Extension of the landscape of a conscious CR
The actual landscape of a CR binds together parts of its instantaneous landscapes during its actual present (cf. Section 1). For a higher CR, the instantaneous landscapes contain perspectives of lower level objects B, not strong or stable enough to be observable in the actual landscape.
A conscious CR is based on higher cortical areas. From the existence of cortico-thalamic links, it follows that a diffuse activation of its agents (in particular because of a fracture in its actual landscape) will activate the reticular formation; roughly, awareness increases. The links from the reticular formation to other areas (here we use the other part of the loop between cortex and reticular formation) transmit their activation in a diffuse way, thus increasing the 'noise'. It allows for an extension of the actual landscape thanks to a process of stochastic resonance which has been experimentally observed (cf. Wiesenfeld & Moss 1995; Levin & Miller 1996; Collins et al. 1996).
For instance, if a clock strikes 4 times, each sound briefly activates auditory receptors, but the record of the sound in Mem lasts longer. Thus, during a short period after the clock stops ringing, we can find what hour it has indicated by recalling not the direct effect of the sounds which have evaporated, but their records.
Let us consider a link from a (cat-)neuron M to an agent A whose perspective is not observable in the actual landscape. A simultaneous activation of M and of A through increasing noise or awareness will lead to a strengthening of the link (Hebb rule). If the activation is large enough, the perspective of the link may become observable in the conscious actual landscape retroactively extended.
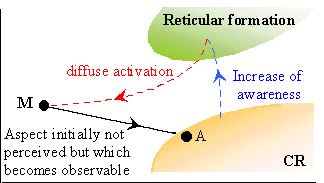
The retroactive extension of the actual landscape relies on this strengthening of perspectives included in some instantaneous landscapes, but initially not strong enough to be retained in the actual landscape. Thus it uncovers weaker internal reflections of the events leading to the fracture, so that its causes might be discovered by the retrospection process.
The processes of prospection and multiple planning of strategies for several steps ahead are operated in virtual extended landscapes, which give an internal representation of how the landscape could evolve taking into account former experiences. The construction relies on several cerebral circuits, in particular those responsible for the links from the CR to the motor cortex (to command effectors) and to frontal areas (for access to the semantics). In particular, it has been proved (Koechlin et al, 1999) that the fronto-polar prefrontal cortex (which is specially well developed in humans compared to other primates) selectively mediates the ability, necessary for multiple planning, to hold goals in mind while exploring and processing secondary goals.
9. Remarks and Complements
9.1. Consciousness and Time
Consciousness is a detector of causality, which gives a selective long term advantage since it allows to react on the causes of a fracture, while a non-conscious CR can only neutralize the effects of a fracture in a purely symptomatic way, under external constraints or as a result of a trial-and-error learning.
With its oscillation between the retrospection and prospection processes, it internalizes the notion of time contrasted to the more stable image of the world assumed by the memory. Past and future are only experienced through their repercussions in the actual present of the conscious CR. A past event Q is not recalled as such, nor even by its more or less stable record M in Mem, but by the perspective of M in the (extended) actual landscape; thus it acquires a coloration depending on the context (e.g., our mood influences our view of a situation). Future is deduced from the construction of virtual extended landscapes in which we project our desires and fears. To be compared with Kant for whom time is the form of our internal state; and with Merleau-Ponty (1945), for whom "la conscience déploie ou constitue le temps" and time "n'est pas un objet de notre savoir, mais une dimension de notre être".
The formation and the extension of the conscious landscape from the instantaneous landscapes during the actual present are exemplified by the confusing experience of a total blank sometimes felt after a brusque wakening. Indeed, the preceding conscious working memory is void, and the formation of the new actual landscape requires the duration of one actual present to bind instantaneous landscapes; aspects coming from the higher levels of the memory are specially long to be activated because the metabolism is slowed, so that the Semantic Memory comes back gradually, through new perceptions which reactivate items in the memory, themselves reactivating other items to which they are linked, up to the restoration of a clear vision of the situation.
9.2. Neurological bases
The model developed in this paper (and already proposed in EV 1992) has been much influenced by the ideas of Edelman (1989) and of Changeux (1983) on consciousness. In particular the study of the neurological bases of primary consciousness by Edelman has given us an experimental support for the processes by which our conscious CRs are characterized.
The model also agrees with the more recent study of Crick (1994) on visual consciousness. Crick assumes the existence of 'treatment units' (unconscious, as in the V1 area) and of 'conscious units'. Each unit has its own form of semi-global representation, its own delays for the treatment and its own access to a short term memory in which the thalamo-cortico-thalamic circuit acts as a reverberating circuit. These properties are exactly the same as those we have attributed to our CRs, the treatment units corresponding to lower CRs, the conscious units to our conscious CRs. The reverberating circuits he considers are those which allow for the extension of the actual landscape and the retrospection and prospection processes in the extended landscape.
9.3. Language and Self-consciousness
Consciousness does not need language. But language allows for more efficient processes, because complex information (higher level cat-neurons) can be stored in the more compact form of a word, and thus more operations can be effected on it. The consequence is the development of a richer 'algebra of mental objects' and of higher order cognitive processes.
A higher conscious CR will form a concept of 'self' if it can analyze its actual landscape as a whole and internalize its processes. For man, this depends heavily on social interactions and communication, through language and education. Self-consciousness relies on a notion of 'continuous identity', which arises from the partial overlapping of successive conscious actual landscapes and their access to the more stable memory. It arises as a construction, initiated by the existence of conscious fractures which reveal the existence of a 'non-self' and, by opposition, lead to the differentiation of 'self'. Whence the "Cogito ergo sum" which signifies that thought, as the conscious perception of a sequence of fractures, makes sense of my existence by contrast with my limitations.
III Illustration: Brief garden scenery on consciousness
Here the above study is illustrated on a concrete example in which the several operations are described in usual language, and then translated in terms of a conscious CR.
10. General example
10.1. Instinctive recording of multiple stimuli
The sight errs. The 'eye' records several images of objects, of spots, of color, of light, of shapes. Simultaneously the mind performs another task, for instance looks for a possible motion indicating the presence of something (a prey for the predator, its predator for an animal, an object of curiosity or study for a man...).
Formation of the landscapes of several lower CRs which receive information on various data while the conscious CR keeps on its actual strategy.
10.2. Unconscious semantic classification of known stimuli, while consciously pursuing the current problem
A spot in the sky appears as a vague form, not very definite. It can be semantically associated with many things:
• undefined without interest (indistinct spot, fly or other insect,...),
• not very distinct nor interesting (very far small bird, flying leaf, airplane),
• more determined and interesting, but still imprecise (distant bird).
In each of these cases, the autonomous classification has been able to slightly activate the concepts of insect, small bird, leaf, more defined bird, plane, but differentially depending on the observer being a cat, a mouse or a man (plane only for the later, leaf or bird for cat or man, bird for mouse only if it is large enough...).
Attributes of the spot are 'stocked' and analyzed in the lower landscapes, which are included in the instantaneous landscapes of the conscious CR but not observable in its actual landscape, which pursues its actual strategy.
Some semantic circuits are more activated, depending on the interest of the observer, whether he looks for any spot, or for a special kind of spots (strategy). In the first case, his attention will converge on any spot causing a fracture. In the second case, only spots similar to the object of his search will arouse his attention
Depending on the long term search strategy, a spot will or will not cause a fracture to the conscious CR.
10.3. The stimuli, or rather their classified representation, become conscious if there is a 'retrospection' on the preceding unconsciously activated items through an internal autonomous circuit
The surge of attention at the time t+f produces a 'consciousness spike', which allows for a synchronous oscillation between the circuits preexcited by the strategy and the circuits which have stored in the memory the several attributes of the spot. The flight of a plane is far off and linear, with a deep sound, the flight of a bird of prey is curved, slow and easy, alternately coming and going away with small cries, the flight of a small bird will be sinuous, hopping, with specific sounds, the flight of an insect is uncertain, near the grass, and so on.
The phenomena may be fugitive, brief awareness, or prolonged, leading to a fracture in the conscious landscape. In this case, a virtual momentarily autonomous internal world is formed, which takes hold of all the capacity of attention, memorization and observation of the observer, with retrospection allowing for multiple comparisons:
• between the spot at t, t+1, t+2, ... t+f,
• between the spot at t and preactivated circuits,
• between the spot at t+1 and preactivated circuits,
• ...............
• between the spot at t+f and preactivated circuits,
• comparisons between the results of the above comparisons and preactivated circuits,
to constitute a moving reflex loop of the type:
|
classification ® comparison ® perceptive memory of the stimuli ® their semantic extensions and corresponding strategies ® classification ® comparison ® .... |
If the spot causes a fracture in the conscious landscape, increased awareness allows for the extension of the landscape, in which the spot will be more accurately analyzed by recovering the records of its several attributes. This analysis depends on a sequence of abduction and prospection processes, each newly recalled attribute leading to new possible causes, and thence to more adequate strategies to respond. The retrospection/abduction and the prospection processes are intertwined, in successive virtual landscapes, as indicated in the following concrete examples.
11. Scenarios
11.1. First step for man (or cat)
"It is a bird, similar to a bird of prey, even if it is small, it might be a small bird with the flight of a bird of prey, some swifts have a light resemblance with a bird of prey, but his flight is too slow for a swift; it is a bird of prey as the one I saw some days ago above the pasture, almost as the one for which I have hidden behind the tree and which was a bird of prey since it has eaten a mouse",
thinks the man or the cat.
The strategy to identify the stimulus first activates the general concept (the most 'typical'), then its attributes are refined: small, with the flight of a bird of prey, whence two 'strategies': swift or bird of prey. A new retrospection in the extended landscape permits to retrieve some data: the flight is slow, which causes a fracture in the virtual landscape corresponding to the choice 'swift'; thus this choice is excluded. It remains to confirm the choice 'bird of prey'. It is done by searching in the episodic memory for instances of the concept 'bird of prey'; this process reactivates a recently activated instance (which has so remained strong enough), and the strategy then realized (to hide behind the tree); and this recalls another property of this instance (a mouse is eaten), which allows for the final identification of a bird of prey.
11.2. New step for the cat
"Now: a bird of prey, but small, is he not smaller than the one seen some days ago? it could then be a swift, and I could eat it; as a swift does not ground, I have better to climb on the tree"
(thinks the cat).
A complementary information 'too small' contradicts the conclusion 'bird of prey'. Whence a new fracture, followed by the same process, leading then to the concept 'swift', and to a choice of adapted strategy.
11.3. For the mouse
"I don't distinguish well"
thinks the mouse which is occupied and does not realize that a spot may be dangerous, except if it is a large spot, very low which obscures the sun and must then be reacted by quickly hiding in a hole; "no problem the spot is not dangerous and it is good to eat seeds. But where is now the spot? Ash! it has become large, I must hide." TOO LATE.
The mouse, with a less developed conscious CR, can only extend its actual landscape on a brief period. Thus the retrospection/abduction is too vague for allowing to recognize the true cause, and she wrongly chooses to pursue her present eating strategy. When the fatal fracture arrives, the strategy of hiding is well selected, but it is too late for realizing it.
12. Bibliography
12.1. General references
Borsuk, K. (1975); Theory of shape; Monografie Mat. 59, Warsow.
Changeux, J.-P. (1983); L'homme neuronal, Fayard, Paris.
Collins, J., Imhoff, T. & Grigg, P. (1996); Noise-enhanced tactile sensation; Nature 383, 770.
Crick, F. (1994); The Astonishing Hypothesis; Macmillan Publishing Company, New York.
Dennett, D. (1990), La stratégie de l'interprète, NRF Gallimard, Paris.
Diers, Y. (1971); Catégories localisables; Thèse, Université Paris VI.
Edelman, G.M. (1989); The remembered Present; Basic Books, New York.
Eilenberg, S. & Mac Lane, S. (1945); General theory of natural equivalences; Trans. AMS 58, pp. 231-294.
Engert, F. & Bonhoeffer, T. (1997); Synapse specificity of long-term potentiation breaks down at short distances; Nature 388, 279-282.
Frey, U. & Morris R. (1997); Synaptic tagging and long-term potentiation; Nature 385, 533-536.
Hebb, D. O. (1949); The organization of behaviour; Wiley, New York.
Josephson, B. (1998), Extendibility of activities and the design of the nervous system: Third Intern. Conf. on Emergence ECHO III; Helsinki; August 1998.
Koechlin, E., Basso, G., Pietrini, P., Penzer, S. & Grafman, J. (1999); The role of the anterior prefrontal cortex in human cognition; Nature 399, 148-151.
Levin, J. & Miller, J. (1996); Broadband neural encoding in the cricket cercal sensory system enhanced by stochastic resonance; Nature 380, 165-168.
Minsky, M. (1986); The society of mind; Simon & Schuster, New York.
Merleau-Ponty, M. (1945); Phénoménologie de la perception; Ed. Gallimard, Paris.
Miltner, W., Braun, C., Arnold, M., Witte, H. & Taub, E. (1999); Nature 397, 434-436.
Niemeyer,P. and Peck, J. (1997); Exploring Java; O'Reilly.
Rodriguez, E., George N., Lachaux J.-P., Martinerie, J. Renault, B. & Varela F. (1999); Perception's shadow: long-distance synchronization of humain brain activity; Nature 397, 430-433.
Usher, M. & Donnelly, N. (1998); Visual synchrony affects binding and segmentation in perception; Nature 394, 179-182.
Wehr, M. & Laurent, G. (1996); Odour encoding by temporal sequences of firing in oscillating neural assemblies; Nature 384, 162-166.
Wiesenfeld, K. & Moss, F. (1995); Stochastic resonance and the benefits of noise...; Nature 373, 33-36.
Wittgenstein, L. (1953); Philosophical Investigations: Blackwell, Oxford.
Zhang, Li, Tao H., Holt C., Harris, W. & Poo M. (1998); A critical window for cooperation and competition among developing retinotectal synapses; Nature 395, 37-43.
12.2. References (EV) to articles of Ehresmann, A. & Vanbremeersch, J.-P.
(1987) Hierarchical Evolutive Systems: A mathematical model for complex systems, Bull. of Math. Bio. 49 (1) (pp. 13-50).
(1989) Modèle d'interaction dynamique entre un système complexe et des agents, Revue Intern. de Systémique 3 (3) (pp. 315-341).
(1990) Hierarchical Evolutive Systems, Proc. 8th International Conference of Cybernetics and Systems (Ed. Manikopoulos), New York (1990), Vol. 1, The NIJT Press, Newark 1990 (pp. 320-327).
(1991) Un modèle pour des systèmes évolutifs avec mémoire, basé sur la théorie des catégories, Revue Intern. de Systémique 5 (1), (pp. 5-25).
(1992) Outils mathématiques utilisés pour modéliser les systèmes complexes, Cahiers Top. and Géom. Diff. Cat. XXXIII-3 (pp. 225-236).
(1992) Semantics and Communication. for Memory Evolutive Systemes, dans Proc. 6th Intern. Conf. on Systems Research (ed. Lasker), University of Windsor.
(1993) Memory Evolutive Systems: An application to an aging theory, in Cybernetics and Systems, Tata McGraw Hill Pub., New Delhi (pp. 190-192).
(1994) Emergence et Téléologie, Prépublication 14-1. Amiens.
(1996) with JLC Chandler: Are interactions between different timescales a characteristic of complexity? Actes du Symposium ECHO, Amiens (162-167).
(1997) Multiplicity Principle and emergence in MES, Journal of Systems Analysis, Modelling, Simulation 26 (pp. 81-117).
(1999) Site Internet: http://perso.wanadoo.fr/vbm-ehr