Ned
Block
Department of Philosophy
NYU
According to The Bell Curve, Black Americans
are genetically inferior to Whites. That's not the only point in Richard
Herrnstein and Charles Murray's book. They also argue that there is something
called "general intelligence" which is measured by IQ tests,
socially important, and 60 percent "heritable" within whites.
(I'll explain heritability below.) But the claim about genetic inferiority
is my target here. It has been subject to wide-ranging criticism since
the book was first published last year. Those criticisms, however, have
missed its deepest flaws. Indeed, the Herrnstein/Murray argument depends
on conceptual confusions that have been tacitly accepted to some degree
by many of the book's sharpest critics.
Let's first be clear about the conclusion itself. In a recent article on "The Real Bell Curve," Charles Murray grumbles about critics, such as Stephen Jay Gould, who read the book as saying that racial differences in IQ are mostly genetic. Murray answers by quoting from the book:
If the reader is now convinced that either the genetic or environmental explanations have won out to the exclusion of the other, we have not done a sufficiently good job of presenting one side or the other. It seems highly likely to us that both genes and environment have something to do with racial differences. What might the mix be? We are resolutely agnostic on that issue; as far as we can determine, the evidence does not yet justify an estimate (311).
In this passage, Herrnstein and Murray are "resolutely
agnostic" about whether bad environment or genetic endowment is more
responsible for the lower IQs of Blacks. But they indicate no agnosticism
at all about whether part of the IQ difference between Blacks and
Whites is genetic; and given their way of thinking about the matter, this
means that they are not at all agnostic about some Black genetic
inferiority.
The Herrnstein-Murray argument for genetic IQ differences is based on two facts: IQ is 60 percent heritable within the White population; and there is a stable, 15-point difference between averages IQs of Whites and Blacks. With IQ largely genetic in Whites, it is natural to conclude - according to Herrnstein and Murray - that the Black-White difference, too, is at least partly genetic. Their argument has more to it; they raise issues about the pattern and the magnitude of the differences that I will get to later. But the most important flaws in the more complex version are fully visible in the simple argument.
Herrnstein's and Murray's argument depends on thinking of the 15-point IQ difference as divisible into a genetic chunk and an environmental chunk. This picture suggests the following three alternatives:
Extreme Environmentalism: Blacks are genetically on a par with whites, so the IQ gap is all environmental.
Extreme Geneticism: Blacks are environmentally on a par with whites, so the IQ gap is all genetic.
The Reasonable View: Blacks are worse off both genetically and environmentally: some of the gap is genetic, some environmental.
Extreme Environmentalism is thought to be excluded by the 60 percent heritability of IQ. Extreme Geneticism is excluded by well-known environmental effects on IQ together with differences between Black and White environments acknowledged by Herrnstein and Murray. So we are left with The Reasonable View - which postulates some Black genetic inferiority.
Notice, however, that the statement of alternatives blots out a crucial possibility: that Blacks are much worse off than Whites environmentally and better off genetically. Allowing this option, we get a different set of alternatives: genetically, Blacks are worse off - or better off - or equal to Whites. I don't say that it is likely that blacks are better off genetically than whites, but it is possible, and--a very important point--what you consider possible affects what you think is an extremist position. Moreover, the critics of Herrnstein and Murray have tended to trip over this possibility. For example, in a New York Times op-ed critique that describes The Bell Curve as "bogus" and "nothing but a racial epithet," Bob Herbert insists that "the overwhelming consensus of experts in the field is that environmental conditions account for most of the disparity when the test results of large groups are compared." In effect, he uses known environmental effects on IQ to argue for a low degree of Black genetic inferiority: in effect, he accepts a version of The Reasonable View. Even Stephen Jay Gould, in his otherwise excellent article in The New Yorker, missteps here. Apparently accepting The Bell Curve's way of conceiving the issue, he complains that Herrnstein and Murray wrongly minimize the large environmental malleability of IQ. He says that they turn "every straw on their side into an oak, while mentioning but downplaying the strong circumstantial case for substantial malleability and little average genetic difference." Gould does not do enough to guard against the natural interpretation of "little average genetic difference" in the context of discussion of The Bell Curve as little average genetic inferiority of blacks. Several critics in The New Republic (October 31, 1994), in turn, wonder about the size of the "genetic component of the black-white difference," thereby buying into the same way of thinking.
If you accept The Bell Curve's way of putting the options, then the idea that environmental differences between blacks and whites are big enough to account for 15 IQ points looks like extremism. But given the actual alternatives - that blacks are genetically on a par with whites, or worse off, or better off - zero genetic difference doesn't seem extremist at all.
But isn't the idea of Black genetic superiority in IQ a desperate and pathetic attempt to exploit a mere logical possibility? Consider a parallel case. Toe number is genetic in sloths and humans, and humans are observed to have five toes whereas (diurnal) sloths are observed to have three. Is there any real possibility that the genetic toe difference between humans and sloths goes in the opposite direction from the observed toe-number gap? It could be that the three-toed sloth evolved six toes, but we observe only three because of a thalidomide-like chemical which has polluted their food during the years in which we have observed them. But this possibility is only worth mentioning as an example of something extremely unlikely. This example suggests a principle that, though never articulated, underlies all of Herrnstein's and Murray's thinking on genes and IQ:
Fundamental Principle: if a characteristic is largely genetic and there is an observed difference in that characteristic between two groups, then there is very likely a genetic difference between the two groups that goes in the same direction as the observed difference.
Applying this principle to the case of IQ: given the substantial heritability of IQ, if East Asians are superior in measured IQ, then, according to the Fundamental Principle, they are highly likely to be genetically superior; and if Blacks are inferior in measured IQ, then they are highly likely to be genetically inferior in IQ.
But while the Fundamental Principle seems intuitively
plausible, it is either irrelevant to the Herrnstein-Murray argument, or
simply false. To see the problem, we need first to understand that the
term "genetic" has two senses. In the next section, I describe
those senses in some detail: to put the point schematically for now, "genetic"
can mean either genetically determined or heritable. Once
that distinction is in place, the problems for the Principle follow. Again,
to put the point schematically for now: if "genetic" is used
to mean genetically determined, then IQ is not genetic, and the
Principle is therefore irrelevant. If "genetic" is used to mean
heritable, then IQ is genetic but the Principle is false. In neither
case, however, does the Principle support the Bell Curve's claim
about genetic differences in IQ.
To understand The Bell Curve's fallacy, we need to distinguish two concepts: the ordinary idea of genetic determination and the scientific concept of heritability, on which all Herrnstein's and Murray's data rely. Genetic determination is a matter of what causes a characteristic: number of toes is genetically determined because our genes cause us to have five toes. Heritability, by contrast, is a matter of what causes differences in a characteristic: heritability of number of toes is a matter of the extent to which genetic differences cause variation in number of toes (that some cats have five toes, and some have six). Heritability is, therefore, defined as a fraction: it is the ratio of genetically caused variation to total variation (including both environmental and genetic variation). Genetic determination, by contrast, is an informal and intuitive notion which lacks quantitative definition, and depends on the idea of a normal environment. A characteristic could be said to be genetically determined if it is coded in and caused by the genes and bound to develop in a normal environment. Consequently, whereas genetic determination in a single person makes sense - my brown hair color is genetically determined - heritability makes sense only relative to a population in which individuals differ from one another - you can't ask "What's the heritability of my IQ?"
For example, the number of fingers on a human hand or toes on a human foot is genetically determined: the genes code for five fingers and toes in almost everyone, and five fingers and toes develop in any normal environment. But the heritability of number of fingers and toes in humans is almost certainly very low. That's because most of the variation in numbers of toes is environmentally caused, often by problems in fetal development. For example, when pregnant women took thalidomide some years ago, many babies had fewer than five fingers and toes. And if we look at numbers of fingers and toes in adults, we find many missing digits as a result of accidents. But genetic coding for six toes is rare in humans (though apparently not in cats). So genetically caused variation appears to be small compared to environmentally caused variation. If someone asks, then, whether numbers of toes is genetic or not, the right answer is: "it depends what you mean by genetic." The number of toes is genetically determined, but heritability is low because genes are not responsible for much of the variation.
Conversely, a characteristic can be highly heritable even if it is not genetically determined. Some years ago when only women wore earrings, the heritability of having an earring was high because differences in whether a person had an earring were "due" to a genetic (chromosomal) difference. Now that earrings are less gender-specific, the heritability of having an earring has no doubt decreased. But neither then nor now was having earrings genetically determined in anything like the manner of having five fingers. The heritability literature is full of cases like this: high measured heritabilities for characteristics whose genetic determination is doubtful. For example, the same methodology that yields 60 percent heritability for IQ also yields 50 percent heritability of academic performance and 40 percent heritability of occupational status. Obviously, occupational status is not genetically determined: genes do not code for working in a printed circuit factory.
More significantly, a child's environment is often a heritable characteristic, strange as this may seem. If degree of musical talent is highly heritable and if variation in the number of the child's music lessons depends on variation in musical talent, then the number of music lessons that a child gets may be heritable, too, despite not being genetically determined. In fact, recent studies of heritabilities of various features of childrens' environments show substantial heritabilities for many environmental features - for example, the "warmth" of the parents' behavior toward the child. Even number of hours of TV watched and number and variety of a childs' toys shows some heritability. If this seems unintelligible, think of it this way: variation in these environmental properties is in part due to variation in heritable characteristics of the child, and so the environmental characteristics themselves are heritable. Readers of The Bell Curve often suppose that a heritable characteristic is one that is passed down in the genes, but this identification is importantly flawed. The number and variety of a child's toys is not passed down in the genes. Heritability is a matter of the causation of differences, not what is "passed down".
I have given examples of traits that are genetically determined but not heritable and, conversely, traits that are heritable but not genetically determined. Do these weird examples have any relevance to the case of IQ? Maybe there is a range of normal cases, of which IQ is an example, for which the oddities that I've pointed to are just irrelevant.
Not so! In fact IQ is a great example of a trait that is highly heritable but not genetically determined. Recall that what makes toe number genetically determined is that having five toes is coded in and caused by the genes so as to develop in any normal environment. By contrast, IQ is enormously affected by normal environmental variation, and in ways that are not well understood. As Herrnstein and Murray concede, children from very low socio-economic status backgrounds who are adopted into high socio-economic status backgrounds have IQs dramatically higher than their parents. The point is underscored by what Herrnstein and Murray call the "Flynn Effect:" IQ has been rising about 3 points every 10 years worldwide. Since World War II, IQ in many countries has gone up 15 points, about the same as the gap separating Blacks and Whites in this country. And in some countries, the rise has been even more dramatic. For example, average IQ in Holland rose 21 points between 1952 and 1982. In a species in which toe number reacted in this way with environment (imagine a centipede-like creature which added toes as it ate more) I doubt that we would think of number of toes as genetically determined.
It is worth emphasizing the solidity of the data about the large IQ increases in Holland. The 21 point increase reported by Flynn is based on comprehensive testing of all Dutch 18 year olds who pass a medical exam (and there has been no change in the pass rate). The test used is Raven's Progressive Matrices, a widely respected "nonverbal test that is an especially good measure of g" (273). Even Richard Lynn, the arch-Jensenist who is the source of much of The Bell Curve's data on race concedes this point. He says "The magnitude of the increase has generally been found to be about three IQ points per decade, making fifteen points over a fifty year period. There have, however, been some larger gains among 18 year-old conscripts in The Netherlands and Belgium amounting to seven IQ points per decade." Lynn also mentions that similar results have been found in France. Herrnstein and Murray concede that "In some countries, the upward drift since World War II has been as much as a point a year for some spans of years" (308). In an area where the facts are often contested, it is notable that this set of facts seems to be accepted by both sides.
One very important conclusion from the Flynn data is that no one understands very much about how environmental variation differentially affects IQ. The cause of the large increases in Holland is simply unknown. Even Herrnstein and Murray concede that "relatively little [of the environmental variation in IQ] can be traced to the shared environments created by families. It is, rather, a set of environmental influences mostly unknown at present, that are experienced by individuals as individuals" (108; emphasis added). Indeed, the crucial factor that has enabled the research that Herrnstein and Murray report to exist at all is the fact that one can measure the heritability of a characteristic without having much of an idea of what the characteristic is. To calculate the heritability of IQ, we do not need to know what IQ tests measure; we need only be able to measure IQ - whatever it is - in various circumstances.
A few additional observations about heritability and IQ will underscore the need for great caution in drawing any inferences about the sources of differences in IQ. A common method for measuring heritability relies on comparisons of the correlations of IQ among one-egg twins raised by their biological parents compared with two-egg twins raised by their biological parents. Suppose your neighbor is one of triplets. One of them is your neighbor's one -egg twin, the other is his two egg (fraternal) twin. Suppose that you can predict the IQ of the one egg twin very reliably from the IQ of your neighbor, but your prediction of the IQ of the two egg twin will be much less reliable. This difference would be an indication of high heritability of IQ because one-egg twins share all their genes whereas two-egg twins normally share half their genes.
Heritability studies of IQ within White populations in the US and northern Europe have tended to yield moderately high heritabilities: Herrnstein's and Murray's 60 percent is a reasonable figure. But it is important to note that no one would do one of these heritability studies in a mixed Black/White population. The reason is straightforward: if you place a pair of Black one-egg twins in different environments "at random," you automatically fail to randomize environments. The Black twins will bring part of their environment with them; they are both Black and will be treated as Black.
Moreover, heritability - unlike genetic determination - can be very different in different populations. For example, the heritability of IQ could be decreased if half the population were chosen at random to receive IQ lowering brain damage: by damaging the brains of some people, you make the environmentally caused variation larger. Or suppose we could make a million clones of Newt Gingrich, raising them in very different environments so there would be some variation in IQ, all environmentally caused. Heritability in that population would be zero because the ratio of genetic variation to total variation is zero if the genetic variation is zero. To take a real example, the heritability of IQ increases throughout childhood into adulthood. One study gives heritability figures of under 20 percent in infancy, about 30 percent in childhood, 50 percent in adolescence, and a bit higher in adult life. Studies of older twins in Sweden report an 80 percent heritability figure for adults by age 50 as compared to a 50 percent heritability for children. One possible reason for the rise in heritability is that although the genetic variation remains the same, environmental variation decreases with age. Children have very different environments; some parents don't speak to their children, others are ever verbally probing and jousting. Adults in industrialized countries, by contrast, are to a greater degree immersed in the same culture (e.g., the same TV programs). With more uniform environments, the heritability goes up. I hope these points remove the temptation (exhibited in The Bell Curve) to think of the heritability of IQ as a constant (like the speed of light). Heritability is a population statistic just like birth rate or number of TVs and can be expected to change with changing circumstances. There is no reason to expect the heritability of IQ in India to be close to the heritability of IQ in Korea.
These issues are pathetically misunderstood by Charles Murray. In a CNN interview reported in The New Republic (January 2, 1995), Murray declared "When I - when we - say 60 percent heritability, it's not 60 percent of the variation. It is 60 percent of the IQ in any given person." Later, he repeated that for the average person, "60 percent of the intelligence comes from heredity" and added that this was true of the "human species," missing the point that heritability makes no sense for an individual and that heritability statistics are population-relative. In a letter to the editor in which Murray complains about being quoted out of context (January 30, 1995), Murray quotes more of what he had said: ". . . your IQ may have been determined overwhelmingly by genes or it may have been - yours personally - or overwhelmingly by environment. That can vary a lot from individual to individual. In the human species as a whole, you have a large genetic component." The Bell Curve itself does not make these embarrassing mistakes. Herrnstein, the late co-author, was a professional on these topics. But the upshot of part of this essay is that the book's main argument depends for some of its persuasive force on a more subtle conflation of heritability and genetic determination. And Murray's confusion serves to underscore just how difficult these concepts can be, even for someone so numerate as Murray.
What's the upshot of the distinction between genetic determination and heritability for the argument of The Bell Curve? Recall the sloth example: Toe number is genetic in sloths and in humans; there is a difference in toe number; so the toe-number difference is genetic. This is a good argument: it strains the imagination to suppose that the genetic toe difference between sloths and humans goes in the opposite direction from the observed toe difference. It is ludicrous to suppose that our genes code for two, despite the five we see at the beach. So in this sense the Herrnstein and Murray argument works for the concept of genetic determination. But the data on genes and IQ are about heritability, not genetic determination.
Is IQ genetically determined as well as heritable? No! As I already pointed out, IQ is very reactive to changes in environments in the normal range. Recall the example of the large rise in Holland. Further, the claim that IQ is genetically determined is not the kind of quantitative claim on which Herrnstein and Murray would want to base their claims about genes and race.
If "genetic" means genetically determined,
then, IQ is not genetic in whites or anyone else (and in any case the issue
is not quantitative), so the Fundamental Principle is irrelevant. If "genetic"
means heritable, however, then IQ is largely genetic (among Whites in the
US at least). But in next section I will show that in this sense of "genetic,"
the argument does not work because the Principle is false.
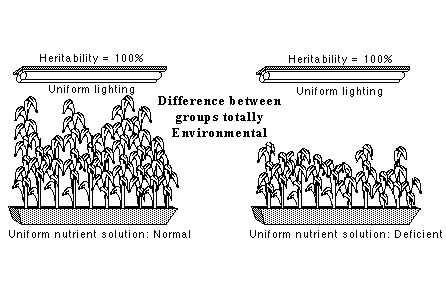
In a 1969 article in the Harvard Educational Review, Arthur Jensen started off the current controversy by arguing from heritability within Whites to genetic differences between Whites and Blacks. Richard Lewontin responded a year later with a graphic illustration of why this is a mistake. Suppose you buy a bag of seed corn from a hardware store. This is not some sort of fancy cloned corn, but ordinary genetically varied corn of the sort that farmers planted long before there was a science of genetics. Grow one handful of it in a carefully controlled environment in which the seeds get uniform illumination and uniform nutrient solution. The corn plants will vary in height, and because the environment is uniform, the heritability of height will be 100 percent. Now take another handful of corn from the same bag, and grow it in a similarly uniform environment but with a uniformly poor nutrient solution. Again, the plants will vary in height, but all will be stunted. Once more the heritability of height is 100 percent. Despite the 100 percent heritabilities of height within each group, the difference in height between the groups is entirely environmentally caused. So we can have total heritability within groups, substantial variation between groups, but no genetic difference between the groups.
The application to race is obvious: heritability is high within Whites. But as Lewontin's example shows, high heritability within groups licenses no conclusion about how to explain differences between groups. - none, in particular, about genetic explanations of the differences. Nor does it dictate the direction of any genetic difference between groups. The stunted corn could have been genetically taller, with the genetic advantage outweighed by the environmental deprivation.
In Lewontin's example, it is assumed that there is no genetic difference between the two groups of corn. But suppose we knew nothing about two groups of people except that they differed by 15 points in IQ and that IQ had some heritability in both, and we had to guess the causes. For all I've said so far, it would make sense to guess that the lower scoring group was disadvantaged both genetically and environmentally. In the next section, I'll show that even this weak principle is wrong. However, the principle has no application to the racial question because we know lots more than nothing: we know that the environment can have huge effects on IQ (e. g., the Flynn Effect of 3 points per decade and the 21 point increase in Holland), and that Blacks are environmentally disadvantaged in a way that has been shown to count. But without being able to measure the effect of being treated as sub-normal, and of an historical legacy of slavery and discrimination, how do we know whether its average effect is sufficient to lower black IQ 15 points, or less than that - or more than that? Given the social importance of this issue, guessing is not appropriate.
Herrnstein and Murray have heard appeals to the legacy of slavery and discrimination. And they have a response which appeals both to the pattern of racial differences and their magnitude.
First, the pattern. They remind us that the Black/White IQ difference is smallest at the lowest socioeconomic levels. And this leads them to ask: "Why, if the Black/White difference is entirely environmental, should the advantage of the 'white' environment compared to the 'black' be greater among the better-off and better- educated blacks and whites? We have not been able to think of a plausible reason. An appeal to the effects of racism to explain ethnic differences also requires explaining why environments poisoned by discrimination and racism for some other groups - against the Chinese or the Jews in some regions of America, for example - have left them with higher scores than the national average" (299).
But these facts are not hard to understand. Blacks and Whites are to some extent separate cultural groups, and there is no reason to think that a measure like socio-economic status means the same thing for every culture. Herrnstein and Murray mention the work of John Ogbu, an anthropologist who has distinguished a number of types of oppressed minorities. A key category is that of "caste-like" minorities who are regarded by themselves and others as inferior, and who, if they are immigrants, are not voluntary immigrants. This category includes the Harijans in India, the Buraku and Koreans in Japan, and the Maori in New Zealand. He distinguishes them from groups like Chinese and Jews who are voluntary immigrants and have a culture of self-respect. If higher socio-economic status Blacks still are to some extent part of a caste-like minority, then they will be at an environmental disadvantage relative to higher socio-economic status Whites. But low status Blacks and Whites are more likely to share a caste background. As Henry Louis Gates, Jr., points out (New Republic, October 31, 1994), affirmative action has had the effect of quadrupling the size of the Black middle class since 1967. Most middle class Blacks have arrived in the middle classes relatively recently, many of them under less than ideal conditions for the development of self-respect. It would be surprising if children of these newly middle-class Blacks were to have fully escaped their caste background in so short a time.
Ogbu notes that where IQ tests have been given, "the children of these caste-like minorities score about 10-15 points . . . lower than dominant group children." He notes further that differences remain "when minority and dominant group members are of similar socioeconomic background." But when "members of a caste-like minority group emigrate to another society, the twin problem of low IQ test scores and low academic achievement appears to disappear. Data suggest that the Buraku who have emigrated to this country do "at least as well at school and the work place" as other Japanese.
As to the magnitude: Herrnstein and Murray calculate that "the average environment of blacks would have to be at the sixth percentile of the distribution of environments among whites. . . for the racial differences to be entirely environmental." And they believe that "differences of this magnitude and pattern are implausible" (299). That is, 94 percent of whites would have to have an environment that is better for the development of IQ than the environment of the average Black - if the 15 point difference is to be explained environmentally. Herrnstein and Murray think this is implausible because when you look at environmental measures - for example, parental income, school quality - you do not find that 94 percent of Whites have a better environment than the average Black. But this calculation ignores the effect of being in Ogbu's category of a caste-like minority. Compare the Dutch 18-year-olds of 1982 with their fathers' cohort, the 18-year- olds of 1952. The difference is entirely environmental despite the probable substantial heritability within each group. Using the same procedures as Herrnstein and Murray, Flynn calculates that 99 percent of the 1982 group had to have a better environment for the development of IQ than the average member of the 1952 group. Given differences of this magnitude among people of a uniform culture who are separated by only a single generation, is it really so implausible that 94 percent of Whites have an environment better than a Black at the 50th percentile?
Environmental differences, then, including the
sort that affect Black Americans, are known to have large effects on IQ.
Moreover, we currently have no way to quantify these effects. So we should
draw no conclusion about the probability of any Black genetic IQ advantage
or disadvantage. As applied to the case of IQ, then, the Fundamental Principle
is false: the combination of high heritability within the White population,
and persistent Black-White differences, does not support a case for genetic
differences.
Earlier, I commented that if we knew nothing at all about two groups except that they differed by 15 points in IQ and that IQ is heritable in both, and we had to guess the causes, it might seem sensible to guess that the lower scoring group was disadvantaged both genetically and environmentally. I have been emphasizing that in the case of Black-White IQ differences, we know much more than "nothing at all." I want now to show that even if we knew nothing, any such guess would be misguided, for reasons that go to the heart of the notion of heritability.
Let's start with an example. Consider a culture in which red-haired children are beaten over the head regularly, but all other children are treated well. This effect will increase the measured heritability of IQ because red-haired identical twins will tend to resemble one another in IQ (because they will both have low IQs) no matter what the social class of the family in which they are raised. The effect of a red-hair gene on red hair is a "direct" genetic effect because the gene affects the color via an internal biochemical process. By contrast, a gene affects a characteristic indirectly by producing a direct effect which interacts with the environment so as to affect the characteristic. In the hypothetical example, the red hair genes affect IQ indirectly. In the case of IQ, no one has any idea how to separate out direct from indirect genetic effects because no one has much of an idea how genes and environment affect IQ. For that reason, we don't know whether or to what extent the roughly 60 percent heritability of IQ found in White populations is indirect heritability as opposed to direct heritability.
The methodology used to measure heritability obscures this ignorance by counting differences in characteristics as caused by genetic differences if there is a genetic difference, even if there is also an environmental difference, thus distorting the ways in which we normally think about causation. For instance, the heritability methodology focuses on the difference between the red hair genes and genes for other hair colors, not on the fact that red-haired children - unlike blond children - are beaten.
Earlier I said that wearing earrings used to be highly heritable because differences were "due" to the XY/XX difference. I put quotes around "due" because it is a by-product of the methodology for measuring heritability to adopt a tacit convention that genes are taken to dominate environment. When virtually only women were wearing earrings, variation in earrings was as much social as genetic, but counted as highly heritable. If there is a genetic difference in the causal chains that lead to different characteristics, the difference counts as genetically caused even if the environmental differences are just as important. If we adopted the opposite convention - concluding from any environmental difference in two causal chains that the difference are environmentally caused - then we could not use current methodology for measuring heritability, because we have no general method of detecting indirect genetic effects using current techniques. Heritabilities using the two different conventions would be radically different if there are substantial indirect genetic effects.
Recall the examples mentioned earlier about the measured heritabilities of such quantities as number of hours of watching TV. No one should suppose that there is variation in genes for watching TV; this is a case of indirect effects. Consider further the fact that no one would do a heritability study on a mixed Black/White population. I mentioned earlier that if you place a pair of Black one-egg twins in different homes, you automatically fail to randomize environments, because the Black twins will bring part of their environment with them; they are both Black and will be treated as Black. This is an indirect genetic effect par excellence. Implicitly, everyone in this field recognizes that, yet more subtle possibilities of indirect effects are typically ignored.
Recall that heritability is defined as a fraction: variation due to genetic differences divided by total variation. The measure of variation that is always used (though alternatives are available) is a statistical quantity known as variance. One factor that raises variance is a positive correlation between genetic and environmental variables. Suppose that children whose genes give them an advantage in musical talent tend to have parents who provide them with an environment conducive to developing that talent - music lessons, concerts, a great CD collection, musical discussion over dinner, etc. Suppose further that other children who have a genetic disadvantage also have an environment that stultifies their musical talents. The correlation between genes and environment will move children towards the extremes of the distribution, increasing the variance in musical skills.
Variance due to gene/environment correlation (gene/environment "covariance") should not be counted in the genetic component of the variance, and there are a variety of methods of separating out such variance. It is common in behavior genetics to distinguish among a number of different types of covariance. The kind just mentioned in which parents provide genes for musical talent and an environment that develops it is called "passive" covariance because it doesn't depend on what the child does. Reactive covariance is a matter of the environment reacting to the child's qualities, as when a school gives extra music classes to children who exhibit musical talent. With active covariance, the child creates a gene-environment correlation, as when a musically talented child practices musical themes in the imagination or pays attention to the musical environment. Passive covariance can be controlled in heritability calculations by attention to adoption studies in which the double-advantage/double disadvantage does not exist. But reactive and active covariance cannot be measured without specific hypotheses about how the environment affects IQ. And as I observed, little is known - as all parties to the disagreements about genetics and IQ agree - about how the environment affects IQ. So distinguishing reactive and active covariance is on the whole beyond the reach of the empirical methods of our era's "behavior genetics," for those methods do not include an understanding of what IQ is - whether it is information-processing capacity, or whether it has more to do with how information-processing capacity is deployed, e.g. whether it is mainly attention - or how the environment affects it.
These points about covariance assume that there are genes for IQ and that these genes may affect the environment so as to produce effects on IQ that are correlated with the ones that the genes themselves produce. But this way of presenting the issue seriously underestimates its significance. For as the red hair example illustrates, indirect genetic effects needn't work through anything that should be thought of as "IQ genes."
Because we don't know much about how variation in environment differentially affects IQ, we can only guess about how variation in genes differentially affects IQ indirectly, via the environment. Suppose that a child's perceived attractiveness and self-confidence strongly affects how adults interact with children in a way that largely accounts for the variation in IQ. Of course, adults could give some children more attention than others without producing IQ differences, but differences might result from variations in adult attention. Suppose further that personal attractiveness and self-confidence are highly heritable. Then we would have an indirect effect par excellence, and such an effect could, for all we know, largely account for the heritability of IQ. Without an understanding of how the environment affects IQ, we simply have no way of determining how much of the variance in IQ is indirect genetic variance of this sort. Of course, if we knew that some specific adult behavior that is triggered by some specific heritable property of children was responsible for a large component of IQ variation, then we could measure that behavior. But there is no theory of intelligence or IQ that would allow us to have any synoptic grip on such factors.
The upshot is that there may be a large component of heritability due to indirect genetic effects, including (but not limited to) gene-environment correlation, that is outside the boundaries of what can be measured given the mainly atheoretical approach available today. Where does the "gene-environment covariance" show up in heritability calculations? Answer: active and reactive effects that we don't know how to measure inevitably are included in the genetic component. This is often regarded by behavior geneticists as perfectly OK. In practice, if researchers were to actually identify an "un-meritocratic" effect such as the red-hair indirect effect mentioned earlier, they would undoubtedly count the variance produced by the effect as covariance rather than genetic variance. But we have no idea how much of the 60 percent of the variance in IQ that is said to be genetic is of this sort. So in practice, covariance due to indirect effects that people know how to measure - at least if it is flagrantly non-meritocratic - is not counted in the heritability; but other indirect effects are counted as genetic. So what counts as genetic variance (inflating heritability) is a matter of value judgments and of what effects we know about. Surely this makes heritability a lousy scientific concept.
In effect, the field has adopted as an axiom that heritability of IQ can be measured by current methods. Without this assumption, the right conclusion would be that since we cannot separate indirect genetic effects (including certain kinds of gene/environment covariance) from pure genetic variance, no heritability estimate can be made. Why does the field adopt this axiom? I cannot help thinking that part of the explanation is that behavior genetics is a young field, struggling for acceptance and funding, and heritability is a flag that attracts attention to it. Let us return to the speculation mentioned above that the 60 percent heritability of IQ (within Whites) is entirely indirect and due to differential treatment of children on the basis of heritable characteristics. Then the direct heritability of IQ would be zero and we would have no reason to think that anything that could be called genes for IQ (e.g., genes for information-processing capacity) vary in the White population, and no reason to look for genetic differences to explain the 15-point difference between Blacks and Whites. Instead, we would have reason to look for differences in the ways adults interact with children to explain the Black-White IQ difference. So indirect heritability suggests an environmental hypothesis about the measured Black-White IQ difference, one that could perhaps be the object of social policy. Are there reasons to expect indirect genetic effects in the Black-White difference? I mentioned the obvious example of genes for skin color above. But there may be less obvious indirect effects as well. There are many more low birth weight Black babies than White babies. Nothing known appears to rule out a genetic explanation. If blacks are more likely to have genes for low birth weight babies, perhaps the effect could be neutralized by diet or by drug intervention in pregnancy. Certainly, no one should think of genes for low birth weight as "IQ genes".
The points I've just made about indirect heritability show why, as I said at the beginning of this section, any inferences from heritability statistics to genetic disadvantage would be misguided. Such inferences seem plausible if we assume that the heritability of IQ within Whites reflects differences in IQ caused by differences in IQ genes. But the points about indirect heritability show that we don't know whether any of the variation within Whites is due to variation in IQ genes. If we have no real grip on the kinds of causal mechanisms that produce the 60 percent heritability within Whites, we can have no confidence in any extrapolation to Blacks.
Let's call a person's genome (his total set of genes) genetically inferior with respect to IQ if that genome yields low IQ in any normal environment. But what is to count as a normal environment? In the example discussed earlier, genes for red hair yield low IQ within environments that are normal in the environment of the hypothetical society, but in environments that we would consider normal, the red hair genes are irrelevant to IQ. What if the heritabilities observed for IQ are a result of indirect effects that can be changed by changing social practice? Then phrases like "genetically inferior in IQ" and "genetic disadvantage in IQ" will only apply to genomes such as that of Down's Syndrome that yield low IQ no matter what the social practices.
The point about indirect heritability also casts doubt on Herrnstein's and Murray's ideas about genetic social stratification among Whites. If the 60 percent heritability does not reflect IQ genes, then there is no reason to suppose that social classes differ at all in IQ genes. Herrnstein and Murray worry about pollution of the gene pool by immigrants and by large numbers of children of low IQ parents. But if the heritability of IQ is mainly indirect, their emphasis on genes is misdirected. If we lived in a culture that damaged the brains of red-haired children, it would be perverse to complain about genetic pollution when large numbers of red-haired immigrants arrived. Instead, we should try to change the social practices that deprive those with certain genes of an equal chance.