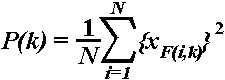
email: jyord@iph.bio.acad.bg
phone: (359) 2-713-37-49
fax: (359) 2-738-469
Time domain analysis of event-related potentials (ERPs) has been conventionally used to study information processing in the brain. In addition, frequency domain analysis has demonstrated that the evoked oscillatory electroencephalographic (EEG) responses in different frequency bands also vary with sensory and cognitive processes (e.g., Basar & Bullock, 1992; Pantev, Elbert, & Lütkenhöner, 1994). The stimulus-related EEG oscillations are defined as the EEG frequency responses or the frequency ERP components and can be extracted from the ERP by appropriate filtering procedures (Basar, 1980; Cook III & Miller, 1992; Farwell, Martinerie, Bashore, Rapp, & Goddard, 1993; Pantev, 1995). It has been assumed that the EEG frequency responses originate from a stimulus-induced reorganization (e.g., frequency stabilization, frequency selective enhancement or damping, and phase-reordering ) of the spontaneous (ongoing) EEG (Sayers, Beagley, & Henshall, 1974; Basar, 1980; Parvin, Torres, & Johnson, 1980) and give raise to the time-domain ERP components (Basar, 1992). In this framework, analysis of both the time and frequency components of the ERP may contribute to a better neurophysiological understanding of stimulus-related brain functioning.
The time domain analysis of ERPs has demonstrated that specific brain mechanisms are activated during the processing of infrequently occurring target stimulus events. Such events produce the parietal endogenous P300 (P3) ERP component, with larger P300 amplitudes obtained under task relevant compared to passive processing conditions (Pritchard, 1981; Johnson, 1986, 1988; Picton, 1992; Polich, 1993), and higher brain processes of attention allocation and memory updating thought to underlie P300 generation (Donchin & Coles, 1988; Polich & Kok, 1995).
In the frequency domain, higher cognitive and associative brain processes have been most consistently correlated with the EEG theta (4-7 Hz) activity (Mizuki, Masotoshi, Isozaki, Nishijima, & Inanaga, 1980; Mizuki, Takii, Nishijima, & Inanaga, 1983; Lang, Lang, Diekmann, & Kornhuber, 1989; Inouye, Shinosaki, Iyama, Matsumoto, & Toi, 1994). Also, it is noteworthy that the EEG theta response has been found to manifest sensitivity to those task variables that are major determinants of the P300 component (Basar-Eroglu, Basar, Demiralp, & Schürmann, 1992; Klimesch, Schimke, & Schwaiger, 1994; Klimesch, 1995). For example, attended and highly relevant stimuli in both the auditory and visual modality have produced in the first 250 ms after stimulation significantly higher theta response amplitudes than task-irrelevant stimuli in passive conditions (Demiralp & Basar, 1992). Further, oddball ERPs have manifested larger theta and delta EEG frequency components compared to passive ERPs (Stampfer & Basar, 1985). Enhanced theta responses with a prolongation up to 500 ms also have been observed for oddball target stimuli but not for passive stimuli (Basar-Eroglu et al., 1992), with a similar prolongation of theta oscillations up to 800 ms found when auditory perceptual difficulty was manipulated (Kolev & Schürmann, 1992). Visual stimuli inducing episodic memory processes have produced event-related theta synchronization that suggests a strong connection between theta activity and memory operations (Klimesch et al., 1994; Klimesch, Doppelmayr, Russeger, & Pachinger, 1996). Taken together, these findings imply that the EEG theta responses in early and late post-stimulus epochs are functionally related to the cognitive processing in task conditions, and second, a relationship may exist between the EEG theta response and P300 ERP component. The aim of the present study was therefore to examine the effect of the "oddball" task processing on the early and late theta ERP components and their association with the P300 typically elicited in this condition (Johnson, 1986).
Despite the overall consistency of the findings from averaged task-related ERPs, however, both single EEG response amplitudes and phase-locking have modified the averaged brain potential amplitude (Woody, 1967; Jervis, Nichols, Johnson, Allen, & Hudson, 1983; McGillem & Aunon, 1987; Ruchkin, 1988; Brandt, Jansen, & Carbonari, 1991). This is an important point to note with regard to functional significance, because single-sweep activity has been shown to reflect specific aspects of stimulus information processing that can be obscured by the averaging procedure (Ford, White, Lim, & Pfefferbaum, 1994; Unsal & Segalowitz, 1995; Yordanova, Kolev, & Basar, 1996). Hence, whether task effects on the evoked theta activity originate from a stronger phase-locking of theta responses to relevant than to irrelevant stimuli, to enhanced single-sweep amplitudes, or to some combination of these factors, is as yet unclear. Further analysis of single-sweep EEG responses in the theta frequency range can provide a more precise evaluation of these task-related effects. To investigate differences between ERPs elicited by passive and active ("oddball") paradigms, the present study assessed EEG theta responses at the level of single sweeps. To analyze single-sweep activity, three parameters were studied: (1) Single-sweep amplitude was used to measure the intensity of the responding system in the theta frequency channel during relevant and irrelevant processing. (2) EEG phase-reordering or stimulus-induced phase-locking was quantified to reflect the stability of the evoked theta patterns independently of amplitude measures. (3) Enhancement relative to prestimulus activity was evaluated to assess event-related variations in the resonant properties of the responding theta system (Basar, 1980, 1992). Specifically, the single-sweep parameters of early and late theta responses and their association with the pre-stimulus theta activity were investigated.
Concerning the relationship between EEG frequency responses and P300
ERP component, many authors agree that the main power of P300 is in the
delta (0.1-4 Hz) range (Duncan-Johnson & Donchin, 1979; Pfefferbaum,
Christensen, Ford, & Kopell, 1986; Stampfer and Basar, 1985). However,
despite the similar functional responsiveness of P300 and EEG theta response
mentioned above, the association between these two time and frequency ERP
components has not been evaluated systematically. In the present study,
the hypothesis of the relationship between P300 and elicited theta
activity (Basar-Eroglu et al., 1992; Intriligator & Polich, 1994; Klimesch
et al., 1994) also was addressed by correlating P300 measures with single
theta response parameters.
Methods
Participants
Fifteen healthy volunteers (nine women, six men) 18-30 years of age
(M = 24.3, SD = 2.1), participated. They were colleagues
and students from the Sofia Medical University; they were right handed,
drug free, and without any history of neurologic, psychiatric disorders,
or hearing problems.
Stimuli and Procedure
The subjects were assessed in a dimly lit, electrically shielded room
and were monitored by means of a closed-loop TV and telephone system. The
auditory stimuli were generated by an IBM-486 computer, filtered, amplified,
and reproduced by a loudspeaker in a free-sound field. All stimuli were
presented at 60 dB SPL, with a duration of 50 ms (rise and fall = 10 ms).
The stimuli were delivered with random interstimulus intervals (3.5 - 6.5
s) in two different task situations. (a) Tone bursts of 800-Hz frequency
(n=50) were presented in a passive listening condition, with participants
instructed only to relax silently. Before the recording session, they were
told that the stimuli would be delivered for testing the technical equipment
and would be of no relevance for them. (b) Auditory target and nontarget
stimuli were presented in an oddball task. During the oddball condition,
100 high and low frequency tones (1200 Hz and 800 Hz) were delivered randomly,
with p = .75 for the high tones and p = .25 for the low tones.
Participants were required to press a button with their dominant hand as
quickly and accurately as possible in response only to the low tones. Reaction
times and error rate were recorded. In both the passive and oddball conditions,
the participants were instructed to keep their eyes closed.
Data Collection and Processing
Electrodes. The EEG data were
recorded with Ag-AgCl disc electrodes placed on midline frontal, central,
and parietal sites (Fz, Cz, and Pz), with linked-mastoids as the reference.
The ground electrode was positioned on the forehead. The electrooculogram
(EOG) was recorded bipolarly with electrodes placed below and at the outer
canthus of the left eye. Electrode impedance did not exceed 8 kOhms.
EEG recording, ADC, and data storage. EEG was amplified with band limits of 0.5 and 70 Hz by means of a Nihon Kohden Electroencephalograph (model EEG-4314F). Stop-band filtering (band limits = 48-52 Hz) was used for eliminating interference. The amplified EEG analog signals were digitized with a sampling frequency of 250 Hz and stored on the computer hard disk, with an epoch length of 1024 ms pre- and 1024 ms poststimulus.
Artifact rejection. The stored raw single sweeps were inspected visually off-line to eliminate EEG segments contaminated with blink, muscular or any other type of artifact activity. Also, any EEG or EOG trial exceeding ±50 µV was excluded from further analysis.
Data Analysis
ERP component analysis in both the time and frequency domains was employed
for the averaged and single-sweep potentials.
Time Domain Analysis
Averaged unfiltered ERPs were obtained to measure
the time-domain components, in particular the P300 component (Figure
1a). The number of artifact-free sweeps averaged for each subject in
each condition was between 35-45 for the passive, and between 22-25 for
the target ERPs.
Analysis of Event-Related Theta Activity
To verify the presence of event-related theta activity
(theta response) three methods were used in addition to single-sweep analysis:
(a) estimation of event-related power in the theta range, (b) calculation
of amplitude-frequency characteristics (AFCs) of averaged ERPs, and (c)
response adaptive digital filtering of averaged potentials.
Event-related theta power. The existence
of event-related responsiveness of the EEG activity in a given frequency
band can be demonstrated by the presence of stimulus-induced power changes.
To study whether power changes in the global (e.g., phase-locked
and nonphase-locked) theta activity occur after stimulus presentation,
the instantaneous power in the 4-7 Hz frequency range was calculated according
to the formula:
Amplitude-frequency characteristics. Another way to verify the presence of time-locked theta responses after stimulation is to compute the AFCs of the averaged ERPs (Basar, 1980; Röschke, Mann, Riemann, Frank, & Fell, 1995). The AFC describes the brain system's transfer properties, e.g., excitability and susceptibility, by revealing resonant as well as salient frequencies. It therefore does not simply represents the spectral power density characterizing the transient signal in the frequency domain but the predicted behaviour of the system (brain) if sinusoidally modulated input signals of defined frequencies were applied as stimulation. As reflecting the amplification in a given frequency channel, the AFC is expressed in relative units. Hence, the presence of a peak in the AFC reveals the resonant frequencies interpreted as the most preferred oscillations of the system during responding to stimulus. To calculate the AFCs, auditory ERPs were first averaged (Figure 1a) and then transformed to the frequency domain by means of one sided Fourier transform (Laplace transform) of the following form (Basar, 1980; Solodovnikov, 1960):
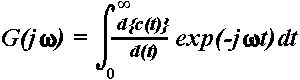
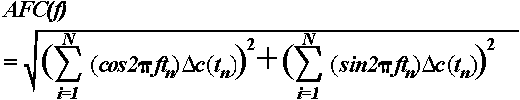 .
.
Here, ![]() c(tn)
represents the first derivative of the transient step response of the
system (in this case, the ERP) at different sampling points tn,
ranging from 1 to N. Although this transform is valid only for linear
systems, it can be applied to nonlinear systems as a first approximation
since errors due to system nonlinearities are smaller than errors resulting
from the length of measurements and rapid transitions in brain activity
(Basar, 1980).
c(tn)
represents the first derivative of the transient step response of the
system (in this case, the ERP) at different sampling points tn,
ranging from 1 to N. Although this transform is valid only for linear
systems, it can be applied to nonlinear systems as a first approximation
since errors due to system nonlinearities are smaller than errors resulting
from the length of measurements and rapid transitions in brain activity
(Basar, 1980).
The AFCs of the averaged ERPs were calculated for
the 0-600 ms (150 data points) epoch after stimulus and then normalized
such that the amplitude ratio at 0 Hz is equal to 1. To enable demonstration
of peaks in different frequency ranges including theta, a log-log presentation
of AFCs was used (Figure 1c).
Digital filtering. To demonstrate further the presence of only phase-locked theta activity, single sweeps in each series were digitally pass-band filtered in the frequency range of 4-7 Hz, and then averaged to reduce the non-phase locked theta components (Kalcher & Pfurtscheller, 1995). Grand average filtered ERPs are illustrated in Figure 1d. To provide a zero phase shift, a modified linear pass-band filter was used, whose weights were based on binomial coefficients (Wastell, 1979). The filter band width was greater than 5% from the total analyzed frequency band, which was experimentally tested to minimize filtering artifacts. To achieve this ratio, the original signals were re-sampled with a sampling frequency of 125 Hz, which introduced no distortion in the signal. The exact half-power frequencies of the digital filter were 3.91 and 7.32 Hz, referred to as 4 and 7 Hz in the text. The length of the filtered single sweep epochs was 2048 ms (-1024, +1024 ms) so that possible edge effects did not alter the analyzed epoch.
Single sweep analysis
As illustrated in Figure 2,
three parameters of the single-sweep theta responses were analyzed for
two time windows, early (0 - 300 ms) and late (300 - 600 ms): (a)
maximal amplitude, (b) phase-locking, and (c) amplitude enhancement against
the prestimulus theta activity.
The maximal peak-to-peak amplitude of the single-sweep theta responses was measured and for each subject, stimulus type, and electrode location the mean value of the maximal peak-to-peak theta amplitude in each of the two time windows was calculated.
For a quantitative evaluation of the phase-locking of single sweeps, a modification of the single-sweep wave identification (SSWI) method was applied (Kolev & Daskalova, 1990; Kolev & Yordanova, 1997; Yordanova & Kolev, 1996). Figure 3 illustrates schematically the analysis procedure, which included the following steps.
First, as shown in Figure 3a, all extrema (minima and maxima) were identified in the filtered (4-7 Hz) single sweeps. Maxima were replaced with +1 and minima with -1. The latency and coded amplitude values of the identified extrema were stored. The detected points along the time axis are presented without the signals (Figure 3b), and the averaged curve is shown in Figure 3c.
Second, as illustrated in Figure 3d, after summing of coded extrema across trials, a histogram of the number of phase-locked single theta waves (SSWI-histogram) can be obtained. To perform data reduction, the analysis epoch was divided into time intervals of 20 ms, which is also within the Nyquist theorem limits for estimation of the frequencies of interest ( where F = maximal frequency in the signal, 7 Hz in this case, and = sampling interval). Thus, for each 20 ms time interval the sum of the identified coded (+1, -1) extrema in the trial set was calculated and the number of the phase-locked waves was determined. The obtained value was represented in the corresponding histogram bar.
Figure 3e presents an example of single theta responses recorded in the passive condition from the Cz lead and visualizes the congruence between sweeps after stimulus. Figure 3f illustrates the corresponding SSWI-histogram. It is clearly observed that in the prestimulus period the phases are random, whereas in the first 600 ms after stimulus presentation the phases are time- and phase-locked. Comparing Figures 3e and 3f demonstrates a dissociation between amplitude and phase-locking for the late (T2) period: although single theta amplitudes in T2 are comparable to those in the pre-stimulus epoch, the responses are random before but phase-locked after stimulation.
Third, quantitative evaluation of single sweep phase-locking was performed. The SSWI-histogram was normalized by dividing the bar values by the number of single sweeps included. The histogram was then rectified (Figure 3h) and the sum of absolute bar values was calculated for the time windows 0-300 and 300-600 ms post-stimulus, thereby giving information about the strength of the single sweep phase-locking in each time window. These measurements were performed for each subject, stimulus type, and electrode site. In addition, to control for effects related to possible changes in frequency, the number of theta waves in each time window was measured at the level of single sweeps and mean individual counts were similarly obtained for each stimulus type and electrode.
To analyze the relationship between pre- and poststimulus amplitude in single theta responses, the maximal peak-to-peak amplitude of each single response was measured and assessed in relation with the maximal peak-to-peak amplitude of the prestimulus theta activity. For each single sweep, the maximal peak-to-peak value of the evoked response was compared with the peak-to-peak value of a theoretical sinusoidal signal having the root-mean-square (RMS) value of the theta EEG activity preceding the stimulus onset (Basar, 1980; Brandt & Jansen, 1991). In such a way, the reference EEG activity was globally characterized for a longer prestimulus period and erroneous detection of maximal peak-to-peak amplitudes occurring randomly in the stimulus-nonrelated epochs was avoided. This comparison is performed by using the enhancement factor, EF (Basar, 1980). As illustrated in Figure 2, for each single sweep filtered in the theta frequency band, the ratio of the maximal response amplitude (max) to the RMS value of the prestimulus EEG amplitude in the epoch -500, 0 ms was calculated according to the formula (Basar, Basar-Eroglu, Rosen, & Schütt, 1984):
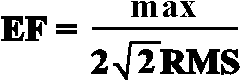
Statistical Analysis
Each of the three parameters of single theta responses (individual
means of the maximal peak-to-peak theta amplitudes, individual means of
enhancement factors, and integral values of the normalized SSWI-histograms)
was subjected to a repeated-measures analysis of variance (ANOVA) with
three within-subjects variables: stimulus (passive vs. oddball),
time window (early vs. late), and electrode (Fz, Cz, and Pz). Individual
mean counts of theta waves were similarly analyzed. The Greenhouse-Geisser
correction was applied to the analyses with repeated measures factor with
three levels (electrode). The original df and probability values
are reported. Amplitude and latency values of the N1, P2,
N2, and P300 components of the ERPs were subjected to Stimulus
x Electrode ANOVAs with repeated measures.
To evaluate the association between prestimulus theta activity and single sweep theta response parameters, correlation analyses for each electrode site were performed between individual rms theta amplitudes of the prestimulus EEG and individual means of single theta response amplitudes, phase-locking, and enhancement factors. The relationship of single sweep parameters and P300-wave amplitude and latency was also assessed using correlational analysis.
Results
Behavioral Data
All subjects performed the oddball task accurately, with the number
of errors being negligible (< 0.5 %) and mean group reaction time equal
to 398 ms (SD = 70.6).
Time Domain ERPs
Figure 1a illustrates the component structure
of auditory ERPs from the passive and oddball stimulus conditions at
Fz, Cz, and Pz. In the passive condition, N1, P2, N2, and a small amplitude
P300-wave were observed, with the P300 most likely originating from the
long and randomly varying inter-stimulus intervals (cf. Picton, 1992; Polich,
Eischen, & Collins, 1994). The usual parieto-central scalp distribution
of the oddball P300-wave was found [electrode, F(2/28) = 8.43, p < .01,
e = 0.71]. Significant differences between passive and oddball ERPs were
observed only for P300 amplitude [stimulus, F(1/14) = 26.5, p < .001],
which was larger for the oddball than for the passive task condition (11.4
µV vs. 3.3 µV).
Theta Power Changes
Figure 1b illustrates that (a) a substantial
power increase occurred for the EEG theta activity after presentation of
each stimulus type; (b) the increase was most prominent at the central
electrode site; (c) the increase lasted longer and was therefore more pronounced
for the 300-600 ms epoch of the target relative to passive ERPs.
Amplitude-Frequency Characteristics
AFCs of grand average ERPs at the three electrode locations are presented
in Figure 1c. The AFCs of both the passive and oddball
ERPs were characterized by a 4-8.5 Hz frequency component peaking at around
6 Hz. AFC components in the slow (1-3.5 Hz) frequency band were also expressed.
A prominent peak was seen at around 10 Hz in response to both types of
stimuli. The figure shows that the values for the slower (delta and theta)
ranges were greater for the target ERPs.
Filtered (4-7 Hz) ERPs
Grand average ERPs filtered in the theta frequency range (4-7 Hz) are
presented in Figure 1d and illustrate that in the
first 250-300 ms pronounced theta responses were elicited by both stimulus
types. Higher amplitudes of the late (after 300 ms) theta responses were
elicited by the oddball stimuli. Significant effects from statistical analyses
are illustrated graphically in Figure 4.
Average theta amplitude (Figure 4a). Early theta responses were larger than the late ones [time window, F(1/14) = 65.1, p < .001]. This effect was much more pronounced for the passive ERPs [stimulus x time window, F(1/14) = 6.13, p = .03], although the stimulus type effect was not significant for either the early or late time window [F(1/14) < 2.7, p > .1]. Maximal amplitudes were observed at the central site [electrode, F(2/28) = 22.1, p < .001, e = 0.98], which was statistically significant only for the early time period [time window x electrode, F(2/28) = 18.4, p < .001, e = 0.74].
Single-sweep theta amplitude (Figure 4b). Early theta response amplitudes were larger than the late ones [time window, F(1/14) = 42.9, p < .001]. The late but not the early responses depended on stimulus type [stimulus x time window, F(1/14) = 7.93, p = .01], and were significantly larger for oddball stimuli [simple stimulus effect for the late theta amplitudes, F(1/14) = 8.84, p = .01]. Single theta amplitudes were maximal at the vertex site [electrode, F(2/28) = 37.9, p < .001, e = 0.96]. This effect, however, was robust only for the early theta amplitudes [time window x electrode, F(2/28) = 13.5, p < .001, e = 0.77].
Phase-locking (Figure 4c). Between-sweep synchronization or phase-locking was significantly stronger for the early relative to the late responses [time window, F(1/14) = 49.2, p < .001]. While phase-locking of the early theta waves did not depend on stimulus type, the phase-locking of the late theta waves was significantly stronger for oddball than for passive stimuli only at Fz and Pz, as indicated by the significant stimulus x time window x electrode interaction [F(2/28) = 7.83, p < .003, e = 0.89], as well as by the site specific post-hoc simple effect tests of the stimulus factor on the late theta phase-locking [Fz: F(1/14) = 5.21, p = .04; Cz: F(1/14) = 2.3, p = 0.88; Pz: F(1/14) = 4.32, p = .05]. These findings are also illustrated in Figure 5 by the SSWI-histograms of one representative subject. They did not result from changes in frequency, because no difference between the passive and oddball condition was found for the counts (number) of theta waves in the late time window at Fz and Pz .
Enhancement factors (Figure 4d). Figure 4d demonstrates that the EFs were significantly higher for the early than for the late theta responses [time window, F(1/14) = 38.8, p < .001]. Only the EFs of the late but not early theta activity were significantly greater to oddball compared with passive stimuli [stimulus x time window, F(1/14) = 10.27, p = .01]. Again, this effect was pronounced over the frontal and parietal locations, with simple stimulus effects being significant only at Fz [F(1/14) = 4.5, p = .05].
Correlation Analyses
Results from testing the relationship between pre-stimulus and evoked
theta activity are summarized in Table 1. Strong and significant correlations
were found between pre-stimulus theta power and theta response amplitudes,
with no associations revealed for the phase-locking measures. The correlational
relationships between single theta response parameters (amplitude, phase-
locking, and enhancement) and the oddball P300 parameters (amplitude and
latency) are presented in Table 2. In sum, these analyzes revealed
that: (a) for early theta responses, across individual subjects,
P300 amplitude was negatively correlated with the phase-locking and enhancement
factors, and P300 latency was positively correlated with the phase-locking;
(b) for late theta responses, P300 amplitude at frontal and parietal sites
was positively correlated with single sweep amplitudes and negatively correlated
with enhancement factors.
|
Table 1. Correlation Coefficients of the Relationship Between Single Theta Response Amplitude and Phase-Locking in Two Time Windows (Early and Late), and RMS Amplitude of Prestimulus Theta Activity (-500 to 0 ms)
* p < .001
Table 2. Correlation Coefficients of the Relationship Between Single Theta Response Parameters in Two Time Windows and P300 Amplitude and Latency of Target ERPs
* p < .05, ** p < .01, *** p < .001 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Discussion
In the present study, a single sweep analysis was applied to auditory theta responses to assess differences between passive and oddball stimulus processing. Previous results from averaged ERPs were confirmed (see below), and single sweep parameters helped to obtain new findings. (a) Both passive and oddball ERPs were characterized by enhanced and strongly phase-locked theta oscillations in the early (0-300 ms) post-stimulus epoch, and by less enhanced and less synchronized theta waves in the later (300-600 ms) epoch. (b) Only the late theta responses differentiated passive from oddball processing by being more enhanced and strongly phase-synchronized to oddball relative to passive stimuli. (c) During target stimulus processing a strong frontal activation was revealed from the phase-locking and enhancement of single theta responses in addition to the parietal brain involvement as reflected also by the P300 component. (d) P300 was strongly associated not only with the concurrent theta responses but also with the theta activity preceding P300 development (prestimulus and elicited immediately after stimulus presentation).
Event-Related Theta Activity
Although in the present study single sweep parameters were used to
describe differences between passive and oddball processing, supplementary
analyses were performed for averaged ERPs to verify the presence of event-related
theta activity and thus substantiate single-sweep analysis results.
(a) Amplitude change (enhancement or damping) of the post-stimulus relative
to the ongoing EEG is a reliable indicator for the presence of event-related
response in a given frequency channel (Basar, 1980; Klimesch et al., 1994;
Kalcher & Pfurtscheller, 1995). Quantification of theta power demonstrated
that after both passive and oddball auditory stimuli, a substantial increase
in theta power occurred within 0-300 ms and lasted longer for the oddball
stimuli (Fig. 1b). These observations will be discussed in detail elsewhere
but they nonetheless provide evidence for the stimulus-related responsiveness
in the theta frequency channel and validate the enhancement factor results
at the level of single sweeps. (b) The amplitude-frequency characteristics
manifested peaks in the theta range (Figure 1c),
which further indicates that oscillatory EEG activity within the frequency
limits of 4-7 Hz was generated and enhanced in relation to stimulus
processing. (c) Prominent oscillatory theta responses were obtained from
the digitally filtered averaged potentials (Figure 1d),
which demonstrates that the stimulus-related theta activity was not only
enhanced but also phase-locked to stimulus since the non-phase-locked responses
were largely attenuated by the averaging. This observation justifies the
independent evaluation of between-sweep synchronization. In general, event-related
EEG activity in the 4-7 Hz frequency band was demonstrated to be present
in the post-stimulus epochs.
Single Theta Responses
In addition to the results from averaged ERPs filtered in the 4-7 Hz
band and amplitude- frequency characteristics that confirmed previous findings
(Basar-Eroglu et al., 1992; Demiralp & Basar, 1992; Schürmann
& Basar, 1994), the independent evaluation of amplitude and phase-locking
at the level of single-sweeps analysis helped to describe more precisely
the EEG theta responses and their association with the ongoing EEG. (a)
It was shown that theta frequency components in the early and late post-stimulus
epochs differ with respect to amplitude, enhancement, and phase-locking
of single sweeps. Hence, the strongly synchronized early theta responses
can be regarded as evoked, and the late theta responses can be referred
to as induced, because of their weaker phase-locking with stimulus (Galambos,
1992; Pantev, 1995). (b) The observed associations between prestimulus
theta activity and single theta responses indicate that whereas response
magnitude depends strongly on the power of the prestimulus theta activity,
the phase-locking of evoked theta waves does not vary with the prestimulus
theta power and appears, therefore, determined mainly by the processing
mechanisms following event presentation. If this hypothesis is accurate,
the phase-locking parameter may prove useful as a sensitive tool for assessing
post-stimulus brain processes. (c) The specific topography patterns and
responsiveness of single theta response parameters to task relevance and
stimulus probability also suggest that single sweep analysis can aid in
assessing neuroelectric correlates of cognitive operations. Indeed, single
sweep parameters revealed differences in task condition processing that
could not be detected at the level of averaged filtered ERPs.
First, single sweep parameters demonstrated that the specific stimulus context was associated only with the late theta responses - an effect that could not be obtained reliably from the averaged filtered (4-7 Hz) ERPs. Hence, higher late theta amplitudes in the averaged ERP observed previously (cf. Stampfer & Basar, 1985; Basar-Eroglu et al., 1992) and in the present study (Figures 1d and 4a) occur because oddball targets produce larger (Figure 4b), better phase-locked (Figure 4c), and more enhanced (Figure 4d) single theta responses within 300-600 ms relative to passive stimuli.
Second, the effects of stimulus type on the single theta responses were most pronounced and significant at the frontal and parietal locations (Figure 4 c d), which indicates that the fronto-parietal associative brain areas might be primarily involved in controlling the late task-related theta oscillations during oddball condition. However, this topography effect again could not be reliably detected in the averaged theta band ERPs (Basar-Eroglu et al., 1992), although it has been observed for the early theta responses to highly predictable attended stimuli in an omitted stimulus paradigm (Demiralp & Basar, 1992). Further, even though P300 amplitude also reflected the stronger parietal involvement for oddball targets in a manner similar to many previous reports with this paradigm (Picton, 1992; Polich & Kok, 1995), single theta response parameters helped to reveal a specific frontal engagement during oddball target processing - effects only hinted at previously when large number of electrodes have been used to map scalp topography effects (Alexander, Porjesz, Bauer, Kuperman, Morzorati, O'Connor, Rohrbaugh, Begleiter, & Polich, 1995). It is not likely that a motor-related activity is responsible for the observed effects because prominent theta responses were elicited also by the passive stimulus which did not require motor responding, and the differences with the oddball stimulus were observed over frontal and parietal but not central areas. Also, it has been previously reported that the late average theta responses to rare auditory targets requiring a mental count were significantly larger than those to passive stimuli (Basar-Eroglu et al., 1992). Finally, on the base of a substantial literature data review, it has been concluded that the correlation of theta activity with motor activity, though important, was full of exceptions and did not fit at all in larger brain animals, and when it occurs is a reflections of a more fundamental correlation with some phases of learning (Miller, 1991, 1994).
Relationships Between the P300 Wave and Single Theta Responses
The P300 component in the present study was significantly larger to
oddball than to passive task conditions (Polich, 1987; Picton, 1992). Likewise,
single sweep parameters of theta activity within 300-600 ms differentiated
passive from oddball target stimuli. Hence, both P300 and late theta responses
appear to be similarly affected by the specific processing of the oddball
target stimulus, which implies that these two brain events might be interrelated.
The possible association between P300 and EEG theta activity has been discussed
in previous studies. Intriligator and Polich (1994) have illustrated that
the relationship between P300 and spontaneous theta activity is sensitive
to target stimulus probability manipulations. Stampfer and Basar (1985)
have demonstrated that changes in the theta frequency band occur after
oddball target presentation, such that they coincide with P300 development.
Basar-Eroglu et al. (1992) and Klimesch et al. (1994) have also suggested
that a linkage between P300 and elicited theta activity exists, because
of their similar responsiveness to signals engaging attention and episodic
memory processes. These data imply that two types of association between
the EEG theta activity and P300 are possible, e.g., functional (or
mediated by relations with common functional mechanisms) and/or direct
(or mediated by the contribution of theta power to P300 expression).
The present study illustrated the similar sensitivity of the P300 and EEG theta responses to oddball target processing. Furthermore, it was demonstrated that individual variation of P300 amplitude at frontal and parietal sites was associated with the amplitude of single theta responses in the late (P300) latency range. This finding shows that the energy of theta frequency ERP component may contribute to individual P300 expression and supports the possibility that event-related theta activity takes part in P300 formation (see also Figure 1 b d). However, across subjects P300 amplitude also correlated with the enhancement factors of the theta responses from the P300 latency range. Since the enhancement factor does not quantify the theta response per se, but rather the relation between pre-and post-stimulus theta activity, P300 amplitude appears to be modulated not only by the concurrent theta oscillations but also by other processes associated with the theta frequency of the EEG. Thus, as indicated by the latter result, the association between the EEG theta activity and P300 may also be functional.
Such a conclusion is further supported by both the present and previous results of the relationship between P300 amplitude and prestimulus/spontaneous theta power (Intriligator & Polich, 1995; Polich, 1996). Furthermore, in this study (Table 2) significant correlations were obtained between P300 (a positive wave occurring about 340 ms post-stimulus) and single theta response phase-locking and enhancement in the latency range preceding P300-wave (0-300 ms). This strong correlation may relate to memory processes because (a) strong synchronization (amplitude enhancement) of theta activity within 375 ms after stimulus has been specifically associated with increased demands toward episodic memory (Klimesch et al., 1994), and (b) the linkage between P300 amplitude and latency measures and working memory processes has also been established (Sanquist, Rohrbaugh, Syndulko, & Lindsley, 1980; Howard & Polich, 1985; Pratt, Michalewski, Barrett, & Starr, 1989; Fabiani, Karis, & Donchin, 1990; Scheffers & Johnson, 1994).
Although the origin of the correlation between P300 and EEG theta frequency needs further elucidation, the present findings demonstrate that individual P300 parameters are associated and modified by the theta-related processes that precede and occur with P300 generation. Thus, in addition to the well documented major contribution of delta frequency component to P300 development and scalp topography (Duncan-Johnson & Donchin, 1979; Stampfer & Basar, 1985; Schürmann, Basar-Eroglu, Kolev, & Basar, 1995; Verleger & Wascher, 1995), the role of stimulus-related theta activity for P300 is strongly emphasized by the present results.
Theoretical Implications
Single theta responses to passive and oddball stimuli can be viewed
in the context of the diffuse theta networks distributed in the brain (Miller,
1991, 1994; Basar, 1992; Lopes da Silva, 1993). In this framework, the
present findings from the three single-sweep parameters (larger amplitude,
stronger phase-locking and enhancement of late theta response to oddball
compared to passive task conditions) might indicate that in the later post-stimulus
period (300-600 ms) a larger number of "traced" or "facilitated" theta
networks can be coactivated simultaneously during task-related than during
passive stimulus processing (Basar, 1980; Pfurtscheller, Steffan, &
Maresch, 1988). Recent theoretical suggestions imply that the scalp theta
EEG activity originates from the interplay between the limbic structures
(hippocampus) and associative (frontal) neocortex of the brain (Miller,
1991, 1994; Basar-Eroglu et al., 1992; Demiralp & Basar, 1992). Because
stimulus type effects reported here were most pronounced at the frontal
and parietal locations, the proposed activation of cortico-hippocampal
feedback loops may be manifested by enhanced and phase-locked single theta
responses.
It may be further hypothesized that two functional aspects of the event-related theta activity may accompany stimulus processing. This assumption is based on the following present and previous findings. (a) Evidence exists for the presence of pronounced early theta response after stimulation. According to the present results, the earlier (0-300 ms) theta response were more enhanced and more strongly synchronized than the later (300-600 ms) ones for both the passive and oddball ERPs. Enhanced theta waves in the early (up to 250 ms) epochs of visual and auditory ERPs also have been described in the studies of Basar (1980), Stampfer & Basar (1985), Schürmann & Basar (1994), and Yordanova et al. (1996). The early theta responses have manifested modality-specific effects because to auditory stimuli they are maximal and best phase-locked over the vertex (Figure 4), while occipital areas were strongly involved for visual stimuli (Klimesch et al., 1994; Basar & Schürmann, 1994). However, it has been also reported that in experiments designed to engage predominantly either semantic or episodic memory, an increase in theta synchronization in the first 375 ms occurred only after presentation of episodic-memory-related stimuli (Klimesch et al., 1994). If early theta synchronization (power enhancement) reflects episodic memory activation, the enhanced and strongly synchronized early theta ERP component may indicate that each presented stimulus entrains some episodic memory processes, with the modality-specific areas being primarily involved. (b) Evidence exists for the presence of task effects on the latency of fronto-parietal theta responses: In both the visual and auditory modality, enhanced theta amplitudes over frontal and parietal regions have been observed for the early but not for the late responses when the stimuli were highly expected and one hundred percent predictable (Demiralp & Basar, 1992). In contrast and as found in the present study, late but not early theta responses at frontal and parietal locations differentiated passive from oddball stimuli. Also, enhanced average theta amplitudes in the late time window (250-500 ms) were obtained for oddball but not for passive or predictable stimuli (Basar-Eroglu et al., 1992). Thus, the task-related effect on fronto-parietal theta responses seems to occur with less delay upon easier condition.
In the context of a parallel sensory-cognitive processing
(Goldman-Rakic, 1988), it may be hypothesized that a "primary" and "modality-related"
theta response is evoked upon stimulus presentation, which develops in
parallel with the concurrent sensory processing such that the incoming
sensory information can be integrated into a general environmental context
(Miller, 1991, 1994). A second order functional engagement or reorganization
of the theta response may occur under the control of the associative (frontal
and parietal) cortical areas if a necessity for further processing is imposed
by a specific task, subjective, or environmental requirement. Further,
this second order theta synchronization may be switched on with a varying
delay or run simultaneously with the early theta processes if the presented
stimuli demand less additional processing by being, for example, highly
probable and easily recognizable. However, further experiments are required
to study whether event-related theta activity reflects specific functional
stages of stimulus and context processing, or a common theta oscillatory
response varies with changing processing demands.
Acknowledgments
This research was supported by the National Scientific
Research Fund at the Ministry of Education, Science, and Technologies,
Sofia, Bulgaria, contracts B-217 and B-225.
We thank Dr. A. Vankov and Dr. T. Demiralp for software
development. Special thanks are due to Dr. John Polich for most helpful
comments and discussions.
Basar, E. (1980). EEG brain dynamics. Relation between EEG and brain evoked potentials. Amsterdam: Elsevier.
Basar, E. (1992). Brain natural frequencies are causal factors for resonances and induced rhythms. In E. Basar & T. H. Bullock (Eds.), Induced rhythms in the brain (pp. 425-467). Boston: Birkhäuser.
Basar, E., Basar-Eroglu, C., Rosen, B., & Schütt, A. (1984). A new approach to endogenous event-related potentials in man: Relation between EEG and P300-wave. International Journal of Neuroscience, 24, 1-21.
Basar, E., & Bullock T.H., Eds. (1992). Induced rhythms in the brain. Boston: Birkhäuser.
Basar, E., & Schürmann, M. (1994). Functional aspects of evoked alpha and theta responses in humans and cats. Occipital recordings in "cross modality" experiments. Biological Cybernetics, 72, 175-183.
Basar-Eroglu, C., Basar, E., Demiralp, T., & Schürmann, M. (1992). P300-response: possible psychophysiological correlates in delta and theta frequency channels. A review. International Journal of Psychophysiology, 13, 161-179.
Brandt, M., Jansen, B., & Carbonari, J. (1991). Pre-stimulus spectral EEG patterns and the visual evoked response. Electroencephalography and Clinical Neurophysiology, 80, 16-20.
Brandt, M., & Jansen, B. (1991). The relationship between prestimulus alpha amplitude and visual evoked potential amplitude. International Journal of Neuroscience, 61, 261-268.
Cook III, E.W., & Miller, G.A. (1992). Digital filtering: Background and tutorial for psychophysiologists. Psychophysiology, 29, 350-367.
Demiralp, T., & Basar, E. (1992). Theta rhythmicities following expected visual and auditory targets. International Journal of Psychophysiology, 13, 147-160.
Donchin, E., & Coles, M.G.H. (1988). Is the P300 component a manifestation of context updating? Behavioral and Brain Sciences, 11, 357-374.
Duncan-Johnson, C.C., & Donchin, E. (1979). The time constant in P300 recording. Psychophysiology, 16, 53-55.
Fabiani, M., Karis, D., & Donchin, E. (1990). Effects of mnemonic strategy manipulation in a Van Restorff paradigm. Electroencephalography and Clinical Neurophysiology, 75, 22-35.
Farwell, L.A., Martinerie, J.M., Bashore, T.R., Rapp, P.E., & Goddard, P.H. (1993). Optimal digital filters for long-latency components of the event-related brain potential. Psychophysiology, 30, 306-315.
Ford, J., White, P., Lim, K., & Pfefferbaum, A. (1994). Schizophrenics have fewer and smaller P300s: A single-trial analysis. Biological Psychiatry, 35, 96-103.
Galambos, R. (1992). A comparison of certain gamma band (40-Hz) brain rhythms in cat and man. In E. Basar & T. H. Bullock (Eds.), Induced rhythms in the brain (pp. 201-216). Boston: Birkhäuser.
Goldman-Rakic, P. (1988). Topography of cognition: Parallel distributed networks in primate association cortex. Annual Reviews in Neuroscience, 11, 137-156.
Howard, L., & Polich, J. (1985). P300 latency and memory span development. Developmental Psychology, 21, 283-289.
Intriligator, J., & Polich, J. (1994). On the relationship between background EEG and the P300 event-related potential. Biological Psychology, 37, 207-218.
Intriligator, J., & Polich, J. (1995). On the relationship between EEG and ERP variability. International Journal of Psychophysiology, 20, 59-74.
Inouye, T., Shinosaki, K., Iyama, A., Matsumoto, Y., & Toi, S. (1994). Moving potential field of frontal midline theta activity during a mental task. Cognitive Brain Research, 2, 87-92.
Jervis, B.W., Nichols, M.J., Johnson, T.E., Allen, E., & Hudson, N.R. (1983). A fundamental investigation of the composition of auditory evoked potentials. IEEE Transaction on Biomedical Engineering, BME-30, 43-49.
Johnson, R. (1986). A triarchic model of P300 amplitude. Psychophysiology, 23, 367-384.
Johnson, R. (1988). The amplitude of the P300 component of the event-related potential: review and synthesis. In P. Ackles, J.R. Jennings, & M.G.H. Coles (Eds.), Advances in psychophysiology: A research annual (Vol. 3, pp. 69-137). Greenwich, CT: JAI Press, Inc.
Kalcher, J., & Pfurtscheller, G. (1995). Discrimination between phase-locked and non-phase-locked event-related EEG activity. Electroencephalography and Clinical Neurophysiology, 94, 381-384.
Klimesch, W. (1995). The P300 and band power in the theta and delta range. Psycoloquy, 95.6.44.memory-brain.5.
Klimesch, W., Schimke, H., & Schwaiger, J. (1994). Episodic and semantic memory: an analysis in the EEG theta and alpha band. Electroencephalography and Clinical Neurophysiology, 91, 428-441.
Klimesch, W., Doppelmayr, M., Russeger, H., & Pachinger, Th. (1996). Theta band power in the human scalp EEG and the encoding of new information. NeuroReport, 7, 1235-1240.
Kolev, V., & Daskalova, M. (1990). Recognition and analysis of single evoked potentials. Proceedings of the North Sea Conference on Biomedical Engineering, Antwerp, Belgium.
Kolev, V., & Schürmann, M. (1992). Event-related prolongation of induced EEG rhythmicities in experiments with a cognitive task. International Journal of Neuroscience, 67, 199-213.
Kolev, V., & Yordanova, J. (1997). Analysis of phase-locking is informative for studying event-related EEG activity. Biological Cybernetics, 76, 229-235.
Lang, M., Lang, W., Diekmann, V., & Kornhuber, H.H. (1989). The frontal theta rhythm indicating motor and cognitive learning. In R. Johnson, Jr., J. Rohrbaugh & R. Parasuraman (Eds.), Current Trends in Event-related Potential Research. Electroenceph. clin. Neurophysiol., Suppl. 40. Amsterdam: Elsevier.
Lopes da Silva (1993). Dynamics of EEG as signals of neuronal populations: models and theoretical considerations. In E. Niedermeyer & F.H. Lopes da Silva (Eds.), Electroencephalography: basic principles, clinical applications, and related fields (3rd ed., pp. 63-77). Baltimore: Williams & Wilkins.
McGillem, C.D., & Aunon, J.I. (1987). Analysis of event-related potentials. In A. Gevins & A. Rmond (Eds.), Methods for analysis of brain electrical and magnetic signals, EEG Handbook (revised series, Vol. 1, pp. 131-169). Amsterdam: Elsevier.
Miller, R. (1991). Cortico-hippocampal interplay and the representation of contexts in the brain. Berlin: Springer.
Miller, R. (1994). Cortico-hippocampal interplay: Synopsis of theory. In C. Pantev, T. Elbert, & B. Ltkenh"ner (Eds.), Oscillatory event-related brain dynamics (NATO ASI Series, Vol. 271, pp. 43-52). New York: Plenum Press.
Mizuki, Y., Masotoshi, T., Isozaki, H., Nishijima, H., & Inanaga, K. (1980). Periodic appearance of theta rhythm in the frontal midline area during performance of a mental task. Electroencephalography and Clinical Neurophysiology, 49, 345-351.
Mizuki, Y., Takii, O., Nishijima, H., & Inanaga, K. (1983). The relationship between the appearance of frontal midline theta activity (FmO) and memory function. Electroencephalography and Clinical Neurophysiology, 56, 56-56.
Pantev, C. (1995). Evoked and induced gamma-band activity of the human cortex. Brain Topography, 7, 321-330.
Pantev, C., Elbert, T., & Ltkenh"ner, B., Eds. (1994). Oscillatory event-related brain dynamics. New York: Plenum Press.
Parvin, C., Torres, F., & Johnson, E. (1980). Synchronization of single evoked response components: Estimation and interrelation of reproducibility measures. In G. Pfurtscheller, P. Busser, F. Lopes da Silva, & H. Petsche (Eds.), Rhythmic EEG activities and cortical functioning (pp. 203-217). Amsterdam: Elsevier.
Pfefferbaum, A., Christensen, C., Ford, J., & Kopell, B.S. (1986). Apparent response incompatibility effects on P3 latency depend on the task. Electroencephalography and Clinical Neurophysiology, 64, 424-437.
Pfurtscheller, G., Steffan, J., & Maresch, H. (1988). ERD mapping and functional topography: temporal and spatial aspects. In G. Pfurtscheller & F.H. Lopes da Silva (Eds.), Functional brain imaging (pp. 117-130). Toronto: Huber.
Picton, T.W. (1992). The P300 wave of the human event-related potential. Journal of Clinical Neurophysiology, 9, 456-479.
Polich, J. (1987). Comparison of P300 from a passive tone sequence paradigm and an active discrimination task. Psychophysiology, 24, 41-46.
Polich, J. (1993). P300 in clinical applications: meaning, method, and measurements. In E. Niedermeyer & F.H. Lopes da Silva (Eds.), Electroencephalography: basic principles, clinical applications, and related fields (3rd ed., pp. 1005-1018). Baltimore: Williams & Wilkins.
Polich, J. (1997). On the relationship between EEG and P300: Individual differences, aging, and ultradian rhythms. International Journal of Psychophysiology, 26, 299-317.
Polich, J., Eischen, S., & Collins, G. (1994). P300 from a single auditory stimulus. Electroencephalography and Clinical Neurophysiology, 92, 253-261.
Polich, J., & Kok, A. (1995). Cognitive and biological determinants of P300: an integrative review. Biological Psychology, 41, 103-146.
Pratt, H., Michalewski, H.J., Barrett, G., & Starr, A. (1989). Brain potentials in a memory-scanning task. I. Modality and task effects on potentials to the probes. Electroencephalography and Clinical Neurophysiology, 72, 407-421.
Pritchard, W.S. (1981). Psychophysiology of the P300. Psychological Bulletin, 89, 506-540.
Röschke, J., Mann, K., Riemann, D., Frank, C., & Fell, J. (1995). Sequential analysis of the brain's transfer properties during consecutive REM episodes. Electroencephalography and Clinical Neurophysiology, 96, 390-397.
Ruchkin, D. (1988). Measurement of event-related potentials: Signal extraction. In T. Picton (Ed.), Human event-related potentials, Handbook of EEG (revised series, Vol. 3, pp. 7-43). Amsterdam: Elsevier.
Sanquist, Th.F., Rohrbaugh, J.W., Syndulko, K., & Lindsley, D.B. (1980). Electrocortical signs of levels of processing: perceptual analysis and recognition memory. Psychophysiology, 17, 568-576.
Sayers, B.McA., Beagley, H.A., & Henshall, W.R. (1974). The mechanism of auditory evoked EEG responses. Nature, 247, 481-483.
Scheffers, M.K., & Johnson, R., Jr. (1994). Recognition memory and search for attended letters: An event-related potential analysis. Journal of Psychophysiology, 8, 328-347.
Schürmann, M., & Basar, E. (1994). Topography of alpha and theta oscillatory responses upon auditory and visual stimuli in humans. Biological Cybernetics, 72, 161-174.
Schürmann, M., Basar-Eroglu, C., Kolev, V., & Basar, E. (1995). A new metric for analyzing single-trial event-related potentials. Application to human visual P300 delta response. Neuroscience Letters, 197, 167-170.
Solodovnikov, V.V. (1960). Introduction to the statistical dynamics of automatic control systems. New York: Dover.
Stampfer, G.H., & Basar, E. (1985). Does frequency analysis lead to better understanding of human event-related potentials. International Journal of Neuroscience, 26, 181-196.
Unsal, A., & Segalowitz, S. (1995). Sources of P300 attenuation after head injury: Single-trial amplitude, latency jitter, and EEG power. Psychophysiology, 32, 249-256.
Verleger, R., & Wascher, E. (1995). Fitting ex-Gauss functions to P3 waveshapes: An attempt at distinguishing between real and apparent changes of P3 latency. Journal of Psychophysiology, 9, 146-158.
Wastell, D.G. (1979). The application of low-pass linear filters to evoked potential data: filtering without phase distortion. Electroencephalography and Clinical Neurophysiology, 46, 355-356.
Woody, C.D. (1967). Characterization of an adaptive filter for the analysis of variable latency neuroelectric signals. Medical and Biological Engineering, 5, 539-553.
Yordanova, J., Kolev, V., & Basar, E. (1996). Evoked brain rhythms are altered markedly in middle-aged subjects: Single-sweep analysis. International Journal of Neuroscience, 85, 155-163.
Yordanova, J., & Kolev, V. (1996). Developmental changes in the
alpha response system in children. Electroencephalography and Clinical
Neurophysiology, 99, 527-538.
(Received June 11, 1996; Accepted February
5, 1997)
Address reprint requests to: Assoc. Prof. Vasil Kolev, Ph. D., Institute of Physiology, Bulgarian Academy of Sciences, Acad. G. Bonchev str., bl. 23, 1113 Sofia, Bulgaria. E-mail: kolev@iph.bio.acad.bg