Mental Health Research Institute, Melbourne, Victoria 3052, Australia
Swinburne Center for
Applied Neuroscience, Hawthorne, Victoria, Melbourne, Australia.
Although we now know much of the structure and functional attributes of the brain, and can understand some of the its mechanisms of information processing, we lack certainty regarding the overall dynamical properties of the brain. That is, we have little idea how the subcomponents of the brain are integrated into a functional whole. Recognising that the problem of the brain's overall integration raises poorly posed but important questions, Churchland (1986) discussed the need for the systematic arrangement of neuroscientific theories within hierarchies, so that reduction of some theories to the level of others can take place.
Two areas requiring such comparative reduction are those of attractor neural networks (ANN) and of mechanisms giving rise to the electroencephalogram (EEG). The ANN field has been largely motivated by the need to reproduce brain-like logical operations. In those studies in which ANN properties are compared to activity patterns of real neurones, the emphasis is generally on comparison to single or multiple unit recordings (Amit 1990). Yet since the EEG is an observable property of large fields of real neurones which are engaged in cortical information processing, the ANN field might benefit from considering the dynamics of the EEG, as a means to cast light on the mechanisms of cognitive processes at larger scales, up to that of the entire brain.
We shall argue that a preliminary integration of neocortical EEG theory with ANN principles is now practicable, and that this unification has consequences for both fields, as well as for studies in single-unit neurophysiology.
To demonstrate the utility of our arguments we present accounts of the simulation of both macroscopic EEG and microscopic cellular interactions, basing both types of simulation upon calculations and measurements of relevant anatomical and physiological parameters. Thus we seek to avoid "explanations" depending on simulations which utilise "free" (arbitrary) parameters. The results of these simulations raise further questions about global control processes in the brain. In our attempts to address these further questions we found we needed to consider not only the dynamics of neuronal interaction at intercellular and global levels, but also the interactions of the cortex with the brain- stem.
We have not attempted an exhaustive review of findings and theory in either the EEG or ANN fields. The EEG models upon which our work has drawn most strongly are those proposed by Freeman (1991) for neocortex, and the global model for EEG developed by Nunez (1989) - works each remarkable for experimental and/or theoretical scope, but at first examination, in mutual contradiction, as we shall describe. For ANN work which exhibits properties of relevance to EEG modelling we have drawn principally on the work of Amit and Tsodyks (1990,1991) and Paresi (1986a,b) - ANN authorities who have aimed their work as much as possible toward meeting the requirements of neurobiology.
To link these disparate accounts together we have added some description of the earlier work of our own group, and the concurrent work of others with related concerns.
In the past 25 years Freeman and his coworkers have systematically developed a model of perceptual processing in the olfactory bulb Freeman 1964, 1972, 1975, 1979, 1987a,b, 1988; Freeman & Skarda1985). In a recent paper (Freeman 1991) this body of information was used to make predictions concerning neocortical dynamics. Similar considerations appear to motivate the ongoing work of this group.
Freeman notes that pyramidal cells occur in multilayered, loosely columnar structures, with recurrent inhibition provided by inhibitory surrounds. Within this extended structure Freeman finds it convenient to recognise a sub-unit configuration called a KII set. KII sets are "lumped" approximations of interactive aggregates of cells. The most common KII set is one of interacting aggregates of excitatory and inhibitory cells, with interactions including all combinations of feedforward and feedback relationships. Interactions between KII sets in neocortex are provided by longer range transverse couplings - the excitatory axonal fields of the pyramidal cells. He predicts that similar properties to those his group has found for neuronal aggregates in the olfactory system, will be found in neocortex.
The first key property upon which his model depends is the lagged time response of dendrites to synaptic input. His estimates indicate that the maximum dendritic depolarisation after input occurs 5.8 msec after action potentials reach the afferent synapses - a delay composed of 1.3 msec. of synaptic delay and 4.5 msec. due to passive electrical properties of the dendritic membrane. In conditions of sufficient excitatory drive to all the neurones in the local cortical network, these delays impose a tendency to oscillation, since excitatory cells interact with surrounding inhibitory cells, which in turn provide recurrent connections to the excitatory cells. The oscillation, he predicts, should have a fundamental frequency of 40+ Hz, imposed by the times required for surges of excitation, recurrent inhibition, disexcitation, and disinhibition, to complete a cycle. These surges of firing will impose a similar cyclicity upon the local field potentials (LFP). Coherence between neuronal firing and the EEG near this frequency has been experimentally demonstrated (Eckhorn et al. 1988; Gray et al. 1989). In contrast, EEG at other frequencies may reflect more complicated interactions, and need not be simply correlated with local cell firing.
The absence of broad-band correlation of EEG with action potentials has long been the grounds upon which the EEG has been trivialised as an epiphenomenon by many physiologists (Stryker 1989). The laying of this trivialisation to rest is a small part of Freeman's concern.
In his 1991 account Freeman does not deal with excitatory-excitatory and inhibitory- inhibitory interactions, although he recognises this deficit, and expects these interactions to complicate the picture considerably. Such interactions would make correlation of 40 Hz ECoG and action potentials rather improbable, unless observations were made in states (such as anaesthesia) in which local activity may be relatively decoupled from its surrounds. This may account in part for the controversy which has surrounded the original reports (Tovee and Rolls 1992).
The second key property Freeman proposes will be found in neocortex, analogous to that measured in the olfactory cortex Eeckman & Freeman 1991) is that described by the asymmetric sigmoid curve. This function relates the normalised pulse probability for an individual neuron to the concurrent EEG wave amplitude. Freeman takes its form to describe indirectly the way the pulse density rises with progressive depolarisation of a small mass of cells in response to afferent bombardment, and predicts that the form of the curve is dependent on the state of nonspecific activation.
An important part of Freeman's theoretical derivation of the exact form of the asymmetric sigmoid curve (Freeman 1979) is embodied in the asymptotic limit of the curve. The limit, designated in Freeman's terminology as Qm, is the maximum pulse density which can be sustained by a group of neurones. Qm itself rises with increasing non-specific cortical activation, and is considered by Freeman to impose a stabilising upper bound on the perturbations induced by other inputs to the KII set. It is notable that the asymmetric sigmoid curve is as of yet a theoretical construct. Freeman's experimental results do not convincingly demonstrate the presence of the state-dependent asymptote, since at least as commonly as an asymptotic limit is apparent, pulse density seems to rise exponentially with rising wave amplitude.
Freeman remarks that while both the asymmetric sigmoid curve and the absolute refractory time of neurones guarantee overall stability, the sharp nonlinearity of neuronal threshold confers a sensitive dependence on both initial conditions and ongoing perturbation. Consequently, both the firing rate and the field potential of any pyramidal cell raised above a critical level of excitation would, according to Freeman, be chaotic. Freeman and Jakubith (1993) have embodied the principles of Freeman (1975, 1979,1991) in two simulations which reproduce the predicted chaotic dynamic properties.
Incontrovertible proof that EEG reflects any simple chaotic process is generally lacking. There are grounds for reservation concerning reports of the dimensionality of EEG from direct measurement. Fundamental difficulties lie in the applicability of estimation algorithms to EEG data because of limitation in the size of data sets, noise contamination, and lack of signal stationarity (Ruelle 1994). These difficulties do appear to be overcome in certain situations in which return-map techniques can be applied to controlled cellular interactions (Schiff et al 1994). Without depending upon problematic direct demonstration in all circumstances, the likelihood that cellular interactions are commonly chaotic is made further plausible by Freeman's argument that the Poisson distribution of cell firing rates, and the 1/f nature of the EEG spectrum associated with this, are incompatible with coupled nonlinear limit-cycle oscillations - the only possible alternative. However, this argument depends on an imprecise use of the term chaos.
While the absence of periodic determinate behaviours suggest that local neuronal interactions may rapidly lose information about their initial conditions, this leaves undecided the autonomous dynamic characteristics of local neuronal interactions in the absence of ongoing perturbation. Since any small pool of neurones is under heavy perturbation by afferents, rapid loss of information on initial conditions is to be expected.
If local neuronal interactions are accepted as chaotic, then is the global system necessarily chaotic? An important class of simulation studies suggest that this must be the case (Kaneko 1990, 1992) These studies concern one-dimensional chaotic maps (that is chaotic numerical subprocesses of considerable generality) which are globally coupled, each to all others. Such coupled maps exhibit global chaos, and appear to escape from the law of large numbers and the central limit theorem. The analogy to masses of coupled neurones is evident (Tsuda 1992, 1994). However, the escape from the law of large numbers does not occur in the presence of noise (Kaneko 1990). We will argue in section 4 that this effect of noise is crucial to the type of dynamics observable at macroscopic scale in the ECoG.
The concept that the EEG is an essentially chaotic process at global scale strikes a strong contrast with a well-developed and also very general model advanced by Nunez (1981, 1984). In this model EEG is treated as a linear wave process, and the dynamics of the entire telencephalon is treated as a problem of mass action of coupled neuron-like elements. In application of mass action laws, the applicability of the theorems of central tendency (central limit theorem and law of large numbers) is implicit, but inescapable.
Nunez places emphasis on a number of factors commonly ignored in models of neuronal interaction. These are:
(a) the significant delays due to axonal conduction
(b) the quantitative predominance of long range interactions mediated by the cortico- cortical fibres.
These two factors are of special significance in large brains, especially the human case.
(c) boundary conditions - the global cortical system is a closed medium for the propagation of travelling waves, implying the possibility of global resonance, and standing waves. The alpha rhythm is considered by Nunez to arise as a global standing wave.
(d) the likelihood that EEG waves will obey a law of linear superposition, since they are based upon a measure (average depolarisation of the cortical cells consequent to afferent bombardment) which he expects to be nearly linear over the relevant operating range - ie, directly proportional to synaptic action density.
Nunez' basic dynamic equations have been solved for spherical, spheroidal and other cases approximating the brain. Principal results include a confirmed prediction (Thatcher et al 1986: Burkitt 1994) of a wave propagation velocity of 7-11 m/sec for human alpha waves, and a number of other findings in qualitative agreement with the theory (Nunez 1981,1994).
A strength of Nunez' work has been the consideration of widely differing classes of solution to his fundamental equations, with variation in the key structural parameters. A weakness has been uncertainty as to exactly what ranges of these parameters actually apply in mammalian brains.
Nunez has recognised explicitly that both local and global interactions may be relevant to the origin of EEG (Nunez 1989
Our group has attempted to bridge the microscopic and global scales by providing a concept which recognises non-linear dynamics at microscopic scale, and emergent linearity of macroscopic electrocortical waves (Wright 1990; Wright et al. 1990a,b; Wright & Sergejew 1991).
A unit structure in the cortex capable of oscillation was postulated - one which was loosely defined, but considered to be a mass of excitatory and inhibitory cells roughly equivalent to a minicolumn. Each oscillator was considered symmetrically and reciprocally coupled to many neighbours, by essentially excitatory couplings. The system variables are the local field potentials associated with each unit oscillator.
For conditions pertaining to the ECoG, such a system can be described in a highly general way by stochastic second-order differential equations, in which the parameters of the state equations (representing the analogs of natural frequencies, damping coefficients, additive and multiplicative couplings of simple harmonic oscillators) are all considered to freely vary to whatever values are needed to fit the behaviour of the system variables. Time-variation of these free parameters thus describes deviations from the simple harmonic case, arising from the extreme non-linearity of the underlying neuronal properties, and the ongoing influence of noisy input from the reticular activating system.
We then assumed that because of the idiosyncrasy, complexity, and noisiness of local cellular interactions, the time-varying parameters were stochastically independent in the large.
It follows (Wright 1990) that despite the extreme nonlinearity of the elements, the macroscopic wave motions at low frequencies in such a system tend to approximate the linear dynamics of coupled simple harmonic oscillators, for which the free parameters are constants. That is, waves obey a superposition principle. The lowest resonant mode frequencies tend to constant values. Energy is equipartitioned among the resonant modes, yielding a "1/f" type of average spectrum. Travelling wave velocities at different frequencies tend to constancy, and are governed essentially by axonal and dendritic delays.
These statistical tendencies hold more closely, the lower the frequency and the longer the wavelength considered. Some of these tendencies reverse at high frequencies, but high frequency activity is greatly attenuated in its contribution to total amplitude of the macroscopic ECoG.
(a) the distribution of damping factors in autoregression (AR) analysis of the ECoG is that expected in a near-equilibrium process, with equipartition of energy among resonant modes. (Wright et al 1990a)
(b) linear inverse filter methods can be applied to the averaged evoked response to determine the impulse giving rise to the evoked response. The forward filter can then be applied in such a way that the likely linearity of the wave process is demonstrated. (Wright et al 1990b)
(c) an equation describing coherence at separated points on a medium traversed by multidirectional linear non-dispersive waves, can be fitted to coherence measures from multichannel ECoG. The distribution of the residuals of fit supports the physical validity of the equation. (Wright and Sergejew 1991)
From the above, it seems reasonable to view interactions among cortical cells as a two- scale phenomenon, with interaction across scales. If we consider the interactions of single neurones in a small locale (ignoring for the moment perturbations from surrounding cells) then the free parameters utilised in a stochastic description cannot be expected to be independent. Thus laws of central tendency may be violated and chaotic and/or limit cycle nonlinear interactions are to be expected. But consider the synaptic activity impinging upon this small locale, arising from the sea of surrounding cells. Linear wave attributes may apply to the macroscopic ECoG fields correlated with the sources of this synaptic bombardment, and the action potential flux in the afferent volleys will reflect the space average of LFPs in all the efferent fields.
The abstract second order oscillator of the stochastic model may be taken to correspond to the KII set of Freeman. The stochastic model can be consistent with Freemans formulation, if interaction among individual cells involves noisily perturbed chaotic activity, particularly if this chaotic activity is of high Lyapunov exponent. The presence of global chaos in the coupled chaotic maps studied Kaneko (1990) may not be relevant in the cortex, when ongoing noisy perturbation is provided by the reticular formation, in conditions of cortical activation. By extension, the linearised, mass action treatment applied by Nunez within his general coupling equations would also be justified, at least as first approximation. But we are left at this point with little idea as to the ways in which local cellular dynamics might overide their merely stochastic participation in the global dynamics - a requirement, surely, since it is a neurophysiological commonplace that individual cells reliably exhibit transient bursts of activity in response to selective and remote stimuli. Indeed, without such a capacity for relatively autonomous activity, how are neurones to exhibit any form of local ANN behaviours?
We therefore turned to simulation studies at both macroscopic and microscopic levels, to see to what extent consistency in such simulations could be maintained, while describing essential experimental data at both levels.
Before simulation could be attempted, an adequate description of connectivity in the cortex had to be provided, and simulations devised which were devoid of arbitrary parameters. Further, the simulations at macroscopic and microscopic levels had to share a consistent parametisation as fully as possible.
To simulate realistic cortical networks, general rules in the form of equations describing the density of interaction of neurocellular components were needed. We have applied a modification of a method initiated by Uttley (1956) to compute the needed connectivities. A detailed treatment is reported in Liley and Wright (1994). The assumptions made in our calculations are:
(i) the basal dendritic system and intracortical axonal trees of a neurone are distributed with average fibre densities which decline exponentially with distance from the cell body, and with spherical symmetry
(ii) the apical dendritic tree can be ignored for calculation of intracortical connectivity since it is involved mainly in cortico-cortical synapses
(iii) only axo-dendritic connections need be accounted for
(iv) an axon and dendrite meeting at random in a "connection space" (defined by the radii of intracortical axons and dendrites) form one synapse. (Equivalently, when a dendrite and axon touch, a synapse forms.)
On the basis of these assumptions, and using the anatomical data and initial calculations of Sholl (1953), and Braitenberg & Schuz (1991), we were able to calculate:
(a) The percent density of synapses in unit cortical volume attributable to cortico- cortical excitatory connections, intracortical excitatory-excitatory, excitatory-inhibitory and inhibitory-excitatory connections, and non-specific afferents.
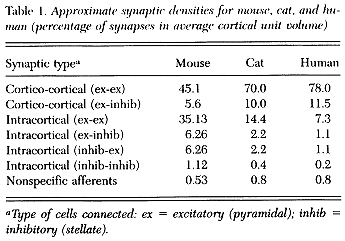
(b) The probability that n synapses are formed between any two cells in the cortex as a function of their separation by a small distance, r.
We have subsequently generalised these calculations so that they can be applied to mammals of variable size, by corrections based on relative cortical cell densities (Wright & Liley 1995). The most recent resultant estimates of synaptic densities for mouse cat and human are given in table 1. Figure 1 shows the probability of synaptic connections between individual cells.
If, as we have assumed, connectivity between individual cells can be appropriately considered stochastic at a local intracortical level, then an important conclusion can be drawn concerning these connections: the connectivity between individual cells is highly asymmetric. Conversely, despite the high asymmetry of individual cell connections, absolute synaptic densities of coupling between pools of cells can be considered roughly symmetric.
These estimates form the basis for the simulations which follow, which thus share a common parametisation of connectivity.
A full account of the macroscopic simulation of human and feline ECoG is given in Wright and Liley (1995), with related aspects elsewhere (Wright et al 1994; Wright and Liley 1994). Each functional unit in the simulation was a "lumped" subsystem, equivalent to a KII set, or the "unit oscillator" of the stochastic model.
The state equations then applied were a specific instantiation of the general equations of Nunez. As such, there was implicit application of the principles of mass action, the law of large numbers and the central limit theorem, as justified by the arguments of section 4.
The state equations described the local field potentials and action potential densities as vector fields over the cortex, with structural parameters introduced to take account of:
(a) Dendritic delays. These were applied to each set of excitatory cells and inhibitory cells within a lumped unit, using a simplified approximation of Freeman's estimates for dendritic transmission delays - ie, a lag of about 5msec to peak dendritic response after afferent bombardment.
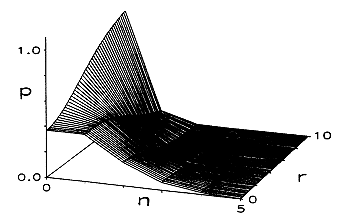 FIGURE 1. A diagrammatic representation of the variation of intracortical synaptic coupling symmetry between two "typical" neocortical pyramidal cells as a function of their intercellular separation. The measure of coupling symmetry (or asymmetry) is the probability that two cells share n presynaptic contacts. Intercellular distances are measured in units of ro, where ro is 31.25 um. (the space constant for the basal dendritic tree). (Liley and Wright 1994) |
(b) Axonal delays, applied to the long range connections. These were equivalent to an action potential propagation velocity of 9 m/sec (again in accord with the figures used by Nunez) and a distribution and maximum range of cortico-cortical connections in accord with anatomical estimates.
(c) Coupling strengths within and between all lumped component groups of inhibitory and excitatory cells which were proportional to the synaptic densities of their connections.
(d) A Gaussian distribution of cell action potential threshold, with respect to somatic membrane potential, or local field potential, giving rise to a sigmoidal pulse-density /LFP relationship for each of the excitatory and inhibitory subcomponents of the lumped units.
(e) Synaptic gain (see below).
Boundary conditions considered for the long-range couplings were the toroidal and zero bound conditions - the former providing a closed geometry sufficient for present purposes, and the zero bound condition being used only for comparison.
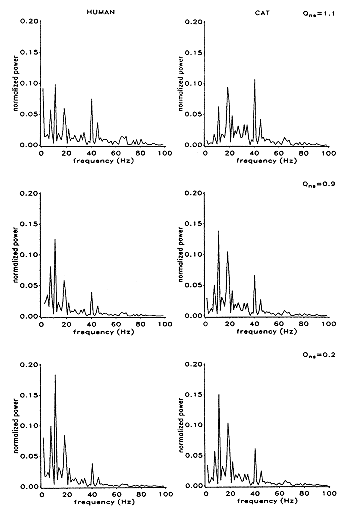 FIGURE 2. Average normalised power spectra (1.22 Hz bins) obtained at steady state from the simulation of human and cat electrocorticogram, for a variety of values of non specific activation. Qns is a measure increasing with nonspecific excitatory tone. (Wright and Liley, 1995) |
Specific sensory input was imitated by input of white noise to a restricted portion of the simulated matrix of lumped cortical elements, and non-specific activation by diffuse input throughout the matrix, weighted by the relative density of non-specific synapses.
The simulation successfully reproduced the following EEG characteristics, some of which are displayed in Figures 2,3 and 4.
(i) The spectral properties of the output included peak densities at the frequencies of the major cerebral rhythms, with a "1/f" spectral envelope (see Figure 2)
(ii) A spectral "shift to the right" with increasing total power occurred as non-specific cortical activation was increased. (also Figure 2)
 FIGURE 3. Phase velocities and attenuation of travelling waves in the cat and human ECoG simulations (Wright and Liley 1995) |
(iii) Steady-state travelling waves with a phase velocity of 5-7 m/sec in the human simulation, and less than 1 m/sec in that of the cat. (Figure 3) were demonstrated. Both velocities are in accord with experimental estimates from the appropriate species Thatcher et al 1986; Burkitt 1994; Lopes da Silva and Storm van Leeuwen 1978; Wright and Sergejew 1991)
(iv) Frequency/wavenumber results (at millimetric scale) revealed an unusual pattern - apparently that of bidirectional and orthogonal standing waves, with a periodic wavenumber spectrum, and wavenumbers independent of the temporal frequency of analysis. This unusual pattern closely matched results from real cat ECoG (Wright et al 1994), and appears to be attributable to reciprocal interaction of each local cortical component with near neighbours. The frequency/wavenumber technique revealed little of the travelling wave activity, most of which lay at wavelengths outside the range estimable by this method. (See Figure 4)
(v) Harmonic responses to strong sinusoidal driving analogous to those generated in cortex during strong visual stimulus by sinusoidal flicker (Wright and Liley 1994a).
The phase velocities and the distributions of power in the frequency/wavenumber results are dependent mainly upon the range and distribution of long range connections. The power spectra show some dependence upon variation in the coupling strengths, but the frequencies of peak power depend principally on dendritic rise and fall times. The 1/f form of the spectral envelope is preserved for all variations of the coupling parameters in which excitatory connections predominate. Thus the model's general properties show relative insensitivity to whatever errors are present in our anatomical estimates.
In these simulations only one parameter - the synaptic gain - was not established a priori from anatomical and physiological data. The value of synaptic gain (g) applied was not arbitrary, although the available means to estimate g was qualitative only. We chose a value of g calculated from the low mean firing rate of cortical cells - approximately 10 spikes/sec, Amit and Tsodyks 1991) - and an estimate of the maximum sustainable firing rate of cortex which does not lead to runaway global excitation. The latter was estimated at only 20 spikes/sec, averaged over all cells in the cortex (Wright and Liley 1995).
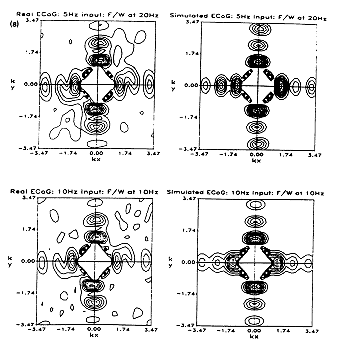
FIGURE 4.
Experimentally observed and simulated frequency/wavenumber plots, for
the cat. During the recordings here analysed, the cat underwent visual
stimulation with a sinusoidally flickering light at the input frequency
indicated, and the simulation was comparably driven with a sinusoidal
"specific sensory" input. As shown, the wavenumbers identified were
invariant with frequency of either the visual driving or temporal
frequency analysed. The same invariant wavenumbers and spatial power
distribution are apparent in "background" ECoG.
Electrode separation 0.86 mm. Each graph shows isocontours (10 to 90%
by 10percentile steps) of normalised power as a function of the
orthogonal components of wavenumber (KX and KY), in radians/mm units.
Power at wavelengths greater in any direction than the dimension of the
array (that is, at wavenumbers near the origin)have been suppressed.
(Wright and Liley 1994, and Wright et al 1994).
|
There are important implications in this choice of g, which will be further considered later. But it can be immediately noted that this choice implies that high firing rates commonly seen for brief periods in single-unit recordings, would, if freely propagated in cortex, lead to runaway excitation, analogous to epileptic discharge. It is further argued in our 1995 paper that the value of g we applied is such that saturation afferent synaptic input to dendritic membranes is just capable of completely depolarising the membrane. Further support for the approximate accuracy of this choice of the value of g arose in our microscopic simulation. Finally, the "shift to the right" of spectral power in the simulated ECoG occurs markedly only as the maximum stable pulse density is closely approached. This spectral shift provides an analog of cortical activation with prominent beta and gamma band activities - a phenomenon which cannot be reproduced in this simulation in association with a realistically low mean firing rate, unless the synaptic gain is high.
We are currently engaged in the study of a cell-by-cell simulation of local cortical dynamics, making use of the "Genesis" simulation package. This work will be reported in detail elsewhere. Its intention is to study in detail the dynamic interactions within locales of individual cells, which were "lumped" under mass action rules, as a component group in the macroscopic simulation.
A thousand or more cortical cells with closely packed cell bodies are represented, with proportions of inhibitory and excitatory cells equivalent to that of real cortex. Intracortical connections are stochastic and highly asymmetrical, in accord with the connectivity rule shown in figure 1, appropriately modified for the pyramidal and stellate cell types.
The parameters of this simulation (connectivities and threshold distributions etc) were chosen so as to correspond to those of the macroscopic simulation as closely as possible. Some features not explicit in the macroscopic simulation are inevitably present, ie:
(i) The location of synaptic inputs on the dendritic tree are modelled, and account taken of the relative distribution of inhibitory and excitatory synapses on the trees.
(ii) Each neurone is in effect a leaky integrate-and-fire process, with 4-8 dendritic compartments, spike threshold discrimination at the soma, and absolute and relative refractory periods.
(iii)Transmitter kinetics, passive dendritic properties and synaptic delay are separately modelled.
(iv) There is no explicit dependence of the model on a synaptic gain term. Instead, this is implicit in the postsynaptic response to each spike, the parameters of which are based on single cell recordings and patch-clamp experiments. Thus no free parameters are involved.
We have so far studied the dynamics of this network under a time-invariant input condition, and differing initial conditions. That is, a uniform and standing direct current stimulus is injected to the dendritic membranes from time zero, so that cellular interactions are enabled to evolve in the net without any time-varying external perturbation. This condition we chose because it is the simplest condition in which to study intrinsic local network properties. The direct current injection can be related to an equivalent average afferent pulse density, as though the network were being bombarded by a steady pulse stream from all cells in the surrounding cortex. The initial conditions of the individual cells can be varied, to obtain some insight into the trajectories followed.
The following properties are apparent:
(i) Two distinct dynamic states emerge, depending on the level of injected current (or external synaptic bombardment) applied during the simulation run. A sharply defined transition between these types of network response occurs at a level of injected current equivalent to a low rate of input of external action potentials. The transition depends upon introducing a level of excitation of the network's component cells which is sufficient to raise the aggregate gain in the intercellular interactions to unity or above.
(ii) At levels of injected current well below this "transition level" the cells remain essentially silent. Just below transition, animations reveal a turbulent pattern of cellular interaction manifested in the membrane potentials. The pattern is sensitive to small changes in the depolarising bias applied to each cell, is associated with low intrinsic firing rates in the network, and time variation of the membrane potentials dies away after initiation of the input bias.
(iii) Above the transition level, runaway excitation in the network supervenes. This terminates in the entry of the network to cyclic activity, in which mean firing rates for the cells are high, and include bursting behaviours. Preliminary results indicate that the network has many different limit cycle, or nearly limit cycle, attractors, and the trajectories followed from different initial conditions can diverge or converge from each other in complicated ways, prior to their ultimate termination in different or similar attractors. The high firing rate states can only be terminated by the application of a strong inhibitory tone to the entire network.
Frames from an animation of network dynamics above the transition level are shown in figure 5.

FIGURE 5. |
We conclude from the results of these simulations that our speculative comments in section 5 are supported in general. The macroscopic and microscopic simulations have been contrived as much as possible so that the one is the generalisation under the principles of mass action of the other. Macroscopic wave motions are near-linear, relatively non-dispersive, and near-equilibrium. A considerable body of experimental data is matched with regard to macroscopic wave motions, and at the microscopic level extremely complicated dynamics, with high sensitivity to initial conditions, are exhibited.
We have not yet been able to define the Lyapunov exponents, or make other dimensional measures of the microscopic dynamics, but we believe these measures could have little meaning without considerable further classification of the basins of attraction present in the state space of any given ensemble of cells.
An effect not considered in our opening sections has become apparent - the sharp transition to excited states of neuronal activity, analogous to a change of phase, in the thermodynamic sense. This property is reminiscent of Freeman's findings in the olfactory bulb, where contrasted dynamics associated with sniffing and resting conditions are apparent (Freeman 1964; Freeman and Skarda 1985).Notably, the sharp transition is a feature of the microscopic model in line with the low maximum firing rate possible without runaway excitation at macroscopic level. Thus the choice of synaptic gain imposed for other reasons in the macroscopic simulation appears independently justified. However, with this choice of synaptic gain the simulations tend to become "stuck" at high firing rates.
Three further points of difference from the parent models from which this work began emerge:
(a) No analog to the state-dependent asymptotic property of Freeman's asymmetric sigmoid function is present in either the macroscopic or microscopic simulations, and therefore no appeal to asymptotic limitation of cell firing rate to stabilise cortical dynamics is applicable within the simulations. Instead, our simple assumption of a Gaussian distribution of action potential threshold for our cell populations imposes a static sigmoidal nonlinearity on wave/pulse relations.
(b) In the macroscopic model, boundary conditions do not exert the great effect on cortical dynamics predicted in most of Nunez' writings, even 'though the simulation reproduces experimental results against which the Nunez model has been previously tested. This appears to be due at least in part to Nunez' assumption that dendritic delays are negligible compared to axonal delays, which is not the case in the simulation. Thus the simulation corresponds to an alternative, and relatively little explored, case within Nunez' general formulation.
(c) The appearance of harmonics in the simulation produced by sinusoidal driving, shows that the macroscopic simulation is significantly nonlinear under perturbation of sufficient magnitude. The stochastic linear model is therefore only approximately applicable, even given free use of the central tendency theorems. The nonlinearity of response is attributable to the static sigmoid gain function, and the near linearity of wave motions depends on the fact that the macroscopic simulation runs in a stable fashion only at low average pulse densities, over which pulse-density/LFP relations are near linear.
We now turn to a brief description of types of dynamics seen in artificial neural networks, which were developed from a different perspective - principally that of the study of memory storage and retrieval.
By "standard models" we mean networks with the following features:
(1) Inputs to each neuron form a linear weighted sum with both positive and negative weight signs permitted.
(2) Outputs of each neuron are two-state (Hopfield 1982) or graded, (Hopfield 1984) with a threshold value for transition.
(3) Connections are bidirectional and symmetric with small lags only.
(4) A system energy (Lyapunov energy) isomorphic with that of an Ising spin-glass can be defined, as a function of the input weights, the current system states, and the unit thresholds.
(5) Elements in the net change state asynchronously, and the system vector follows a trajectory to a point attractor- a static final state. At the attractor Lyapunov energy is at a local minimum.
(6) Initial states leading to a common point attractor are thus classified identically by their shared basin of attraction. Since the trajectories to each point attractor converge, they are not chaotic, by definition.
(7)In the presence of high asymmetry of couplings, an energy is not defined, and system convergence to a point attractor does not take place (Hopfield & Tank 1986).
Amit (1990) draws attention to the limitations of standard models in so far as they may be considered models of real neuronal cooperativity. They are generally applied with the assumption that a network of 104 elements or so is about the size of a realistic pool of completely interconnected neurones in the cortex. Their dynamic properties are rather insensitive to symmetry of coupling strengths, until a point is reached at which the positions in state space of the basins of attraction are not fixed, but are instead functions of the current state.
It may be that mobility of basins of attraction is crucial to the problem of storing and retrieving temporal sequences of inputs since this property confers the ability to output a sequence of different patterns of action potential, rather than the single fixed pattern of activity provided at convergence to a point attractor in a standard network(Caianiello et al. 1967; Amari 1972; Little & Shaw 1975; Peretto & Niez 1986; Nebenzahl 1987; Dehaene et al. 1987).
Dynamic properties of trajectories in the state space of asymmetric nets can include chaotic and limitcycle behaviours. These offer interesting features when asymmetric nets are considered in connection with Hebbian learning (Parisi 1986a,b; Hopfield & Tank 1986). In the presence of a "palimpsest" type of continuously operating synaptic modification, in which recurrent activation increases synaptic weight, while disuse reduces the weight, learning and forgetting occurs depending on the class of dynamics in which neurones are engaged (Parisi 1986a; Nadal et al. 1986). Chaotic dynamics in some synapses prevent storage while other synapses engaged in activity of a limit cycle type store memory.
Nebenzahl (1987) and Dehaene et al. (1987) further consider the case of symmetric couplings of two types between neurones - fast synapses, and those which begin to operate after a delay. This also confers mobility of the attractor basins, with potential for temporal sequencing of the network's behaviour. The associated energy function (or "landscape") is said to be adiabatic (ie., slowly changing).
A neural design advanced by Amit and Tsodyks (1990, 1991) is composed of excitatory cells with inhibitory surrounds. The cells have an absolute refractory period and continuous membrane dynamics with realistic decay constants and rise times. The network is immersed in random continuous afferent input, as if from surrounding cell activity, with short bursts of nonrandom input as the signal to be classified.
These features lead to continuous and coherent intra-network dendritic potentials, usually below action potential threshold, with spikes mainly emitted because of the noisy continuous afferent. Firing rates remain low, but specific neurones spike more rapidly when the state vector is centred on an attractor.
Because of the ongoing noise the attractor dynamics cannot be classified as chaotic, limit cycle, or point attractor; each attractor basin corresponding only to a nonergodic domain.
Amit and coworkers (Griniasty et al. 1993), have shown that their symmetric networks can convert temporal correlations between stimuli to spatial correlations between attractors, paralleling the experimental findings of Miyashita and coworkers (Miyashita & Chang 1988; Miyashita 1988; Sakai & Miyashita 1991). The model can, in principle, be applied in conditions of coupling asymmetry (Amit, personal communication), but it is unclear at what level of asymmetry the definition of basins of attraction would be drastically affected.
Our macroscopic and microscopic simulations of ECoG and local field potential properties can, to a considerable extent, also be understood as special cases of ANN dynamics, as outlined above.
Since cell-to-cell connections appear to be highly asymmetric, the energy concepts applicable in standard models do not apply in our microscopic simulation. The Amit and Tsodyks model is not directly comparable, since this model operates only at low firing rates, and has not been tested at such levels of asymmetry. However, there appears to be a good match with regard to the appearance of coherent fluctuations of local field potentials. The Amit and Tsodyks model appears to be equivalent to a standard model ANN, under noisy perturbation, but with dendritic properties derived from physiological experiment as in our microscopic simulation. The appearance of coherent dendritic potential fluctuations at low firing rates may therefore reflect the way in which dendritic potentials are transduced into action potentials, and depend little on specific connectivities.
It is unclear whether the success of Griniasty et al (1993) in using Hebbian learning to reproduce the experimental findings of Miyashita et al (cited above), could be reproduced in our simulation. The principle source of uncertainty arises from the asymmetry of connections involved in the microscopic simulation. It may be, of course, that the cells involved in the Miyashita experiments share fairly symmetrical couplings, but we know of no reason to believe (or disbelieve) this is the case.
On the other hand, the types of Hebbian learning envisaged by Parisi (1986 a,b) and Nadal et al. (1986) (see section 7.2) appear applicable. In our simulation the combination of extreme divergence and asymmetry of couplings implies the necessary existence of many closed (and thus potentially self-exciting) loops capable of generating limit cycles when the overall firing rate is high, and thus creating basins of attraction.
Our microscopic simulation seems, therefore, to approximate the behaviours of certain asymmetric nets with potential to classify, store, and recall temporal sequences - but to also share properties with one low firing rate model, which describes the behaviour of neural networks surrounded in a sea of concurrent input from the rest of the brain.
For scales greater than the macrocolumn, up to the entire closed extent of the cortex, average couplings can be more appropriately considered symmetrical, as indicated by the general success of our symmetrically coupled macroscopic simulation. Therefore, concepts of system energy are applicable.
The coupled stochastic oscillator model implies, at face value, that a very simple sort of basin of attraction governs the trajectories followed in the cortical state space. Since all long range couplings are excitatory (positive) couplings, this implies a basin of attraction with a single energy minimum, analogous to the case of a ferromagnet in spin-glass theory. If that were so the trajectories followed would be defined simply by the principle of least action. This interpretation does not take account of complications in the form of the energy landscape which might emerge consequent to interactions among the stochastic parameters, each of which was assumed effectively stochastically independent of the others.
Moreover, this "face value" interpretation is belied by the further properties which emerge from the simulations. These are:
(a) The sharp transition from low firing rate to high firing rate dynamics. The appearance of patches of excited activity in the cortex would violate the assumption of independence of parameters in the stochastic model.
(b) The requirement, imposed by this sharp transition, for a stabilisation mechanism to prevent runaway excitation.
These complicating effects have the following implications.
The presence of sharp transitions to rapid firing rates implies that, given sufficient excitement, small pools of cells can temporarily assume the capacity to drive activity elsewhere in the brain. During this state, the involved neurones would be readily and preferentially identified by single-cell recording techniques. Since complicated high firing rate dynamics at specific locii are associated in single cell studies with very specific stimulus or response properties, this appears to be a strength of the simulations.
Conversely, macroscopic scale events must interact with the single unit events, by virtue of the space averaging over large neuronal fields implicit in the determination of the synaptic input to small pools of neurones. This seems to offer a plausible explanation for the widely established association of temporal and topographic EEG with cognitive states (Gevins et al 1983; Picton and Hillyard 1988).
If this interpretation is correct, then local areas of highly excited cells must be capable of highly perturbing the global brain state. The sharp transitions of "phase" about unity aggregate gain may be expected to confer a sensitive dependence upon initial conditions to the ECoG time-series, even though mass action principles apply in the large, and whether or not attractor dynamics are truly chaotic, above or below this transition level of excitation. Highly erratic perturbation about a state approaching equipartition of energy among resonant modes is apparent in activated ECoG ( Wright et al 1990a).
As they are presently formulated, both classes of simulation undergo transition to runaway excitation, and stay there, under all but minimal perturbation. In the case of the macroscopic simulation it is easy to compare this runaway excitation to epilepsy, albeit in a very crude way. Indeed, the need for a low epileptic threshold formed part of our argument in arriving at a value for synaptic gain. But in the microscopic simulation the high firing rate states appear to correspond to attractor states necessary for functional classification of inputs. It is probably impossible to appeal to any local inhibitory mechanisms which might stabilise this runaway excitation, since once local excitation has become sufficiently intense to activate any local control, long range spread of excitation would have already occurred.
Similar instability was encountered in the now classic work of Wilson and Cowan (1973). These workers found that to achieve global stability in a model generically similar to our simulations, they required excitatory-inhibitory interactions to occur at longer range than excitatory-excitatory interactions. This is a condition which cannot be met by a model concerned with the cortex alone. The requirement for stability arrived at by Wilson and Cowan means that large rings of inhibition must be generated around all excited cortical zones - a requirement which appears possible only if cortical-subcortical interactions are brought into play.
There is no shortage of candidate pathways for such control. The cortico-striatal- thalamic pathways, and the long catecholeaminergic and indolaminergic fibres to cortex suggest themselves, but a detailed consideration of the possible pathways of interaction is beyond the scope of this paper.
Cortical-subcortical interaction is generally polysynaptic, topographically organised in parts, and diffuse in others. All delays in the total cortico-fugal and centrifugal extents of the interaction are likely to be much greater than cortico-cortical delays. Therefore rapid control may be expected to involve relatively fast pathways of cortical-subcortical interaction. Perhaps local changes in intracortical inhibitory processes more complex than those introduced to our simulations so far, also play a part, albeit a subsidiary part.
It remains as a large future modelling exercise to determine whether such interactions would be sufficient to retain overall stability, or whether this requirement exposes a flaw in our conception. We have made a beginning by developing a two-cortical-layer version of our macroscopic simulation, and introducing the actions of the neuromodulators noradrenaline and dopamine. These neuromodulators appear to be excitatory in the deeper cortical layers, and inhibitory in the superficial layers (reviewed by Silberstein 1995). We have shown that under this differential excitatory and inhibitory drive, the simulation can exhibit global stability, while local and transient firing rates in the deeper layers transcend the maximum global average compatible with stability - that is patches of cortex can pass the "transition level" without global runaway excitation inevitably supervening.
Results of these two-layer simulations will also be reported elsewhere. An incidental implication of these findings is that neuronal dynamics may be included in the class of type IV cellular automata, thus exploiting the computational potential of controlled phase transitions (Langton et al 1992).
If cortical global stability is eventually proven to depend on cortical-subcortical interactions, then a further mechanism of cerebral self-control, analogous in character to ANN findings, is implicit in the stabilising process. Cortical-subcortical-cortical circuits can be considered equivalent to slow synapses within cortex and could thus act as a form of adiabatic control of the energy landscape. This control would operate on the slope at all points on the global energy basin, and thus partially control the state trajectory for the entire brain.
We have sketched an outline of the possible organisation of brain dynamics, and have indicated that while events at cellular and global scales may be studied to some degree in isolation, the rules by which the brain constitutes a cognitive engine may depend on both local and global network properties, with interactions across scale. This sketch explains basic wave properties of the EEG, and is partially unified with some ANN principles, which themselves appear to offer an account for important aspects of cortical information processing and storage. The future study of learning rules within the simulations we have presented appears practicable.
Our outline appears sufficiently precise to be subject to critical test, as increasingly accurate physiological measurements are made, and the simulations are developed to higher levels of anatomical and physiological veracity. As instances:
(1) in the macroscopic simulation, as cortico-cortical connectivity is modelled with greater anatomical realism, then the predicted estimates of wave velocities and frequency/wavenumber spectra should increasingly approximate more precise physiological estimates.
(2) in appropriate specific instances of the microscopic simulation, the mean firing rates, distributions, and time-series characteristics of individual units should approach those seen in specific single-cell experiments. However, to meet this test, both an adequate model of the self-stabilising interaction with subcortical systems, and much better definition of variations in cortical coupling symmetries may be essential prerequisites.
The simulations we have presented lack detail. Most seriously, they lack specification of the pathways, distributions, and relevant time-constants for the stabilising interactions with subcortical structures upon which their plausibility as genuine models of cortical dynamics depends. We have made little attempt to describe the influences of organisation in depth within the cortex. No regard has been given to the macroscopic and microscopic inhomogeneities of cortico-cortical and intracortical connectivity, nor the effects of rhythmic subcortical processes and resonances between cortex and subcortex, which undoubtably contribute to the EEG (Steriade et al 1990). We have not considered the effects of neuromodulators in any detail, nor any complications of synaptic transmission such as hyperpolarising afterpotentials, nor cortical cellular specificity greater than that of two cell types - pyramidal and stellate. But it would appear that none of these factors, nor many others, are essential to a parsimonious treatment of general dynamic principles.
It also appears likely that the principles we have outlined apply only under certain further restrictions, eg, to waking states. A key point in our argument has been the conclusion that microscopic chaos (in the broadest sense) is compatible with the operation of mass action effects at macroscopic level. This argument depended on the introduction of spatially and temporally noisy input from the reticular formation. It may be that the withdrawal of this input permits the global chaos described in coupled chaotic maps by Kaneko (1990) to emerge.
While the most important reservation on the completeness of this outline of dynamic principles appears to us to lie with specification of the cortical/subcortical interactions, it does appear that such interactions might prove relatively simple to model, for the very reason that to sustain low average firing rate stability, the return pathways to the cortex must exert fast, and widespread, effects. Whether simple or complicated, the effect such control might have on the competitive evolution of dynamic patterns of activity in the cortex should be very rich.
Amari, S. (1972). Learning patterns and pattern sequences by self-organising nets of threshold elements. IEEE Transactions on Computers 21: 1197-1206.
Amit, D.J. (1990). Modelling brain function. The world of attractor neural networks. New York, London, Cambridge University Press.
Amit, D.J. & Tsodyks, M.V. (1990). Attractor neural networks with biological probe records. Network 1: 381-405.
Amit, D.J. & Tsodyks, M.V. (1991). Quantitative study of attractor neural networks retrieving at low spike rates: II. Low rate retrieval in symmetric networks. Network 2: 275-294.
Braitenberg, V. & Schuz, A. (1991). Anatomy of the Cortex: Statistics and Geometry. Berlin, Springer-Verlag.
Burkitt, G. (1994). Steady-state visually evoked potentials and travelling waves. PhD dissertation. Swinburne University of Technology, Melbourne, Australia.
Caianiello, E.R., De Luca, A. & Ricciardi, L.M. (1967). Reverberations and control of neural networks. Kybernetik 4: 10-18.
Churchland, P.S. (1986). Neurophilosophy - toward a unified science of the mind-brain. Cambridge MA, MIT Press.
Dehaene, S., Changeux, J.P. & Nadal, J.P. (1987). Neural networks that learn temporal sequences by selection. Proceedings of the National Academy of Science 84: 2727-2731.
Eckhorn, R., Bauer, B., Jordan, W., Brosch, M., Kruse, W., Munk, M. & Reitboeck, H.J. (1988). Coherent oscillation: a mechanism of feature linking in visual cortex? Biological Cybernetics 60: 121-130.
Eeckman, F.H. & Freeman, W.J. (1991). Asymmetric sigmoid nonlinearity in the rat olfactory system. Brain Research 557: 13-21.
Freeman, W.J. (1964). A linear distributed feedback model for prepyriform cortex. Experimental Neurology 10: 525-547.
Freeman, W.J. (1972). Measurement of open-loop responses to electrical stimulation in olfactory bulb of cat. Journal of Neurophysiology 35: 745-761.
Freeman, W.J. (1975). Mass action in the nervous system. New York, Academic Press.
Freeman, W.J. (1979). Nonlinear gain mediation of cortical stimulus response relations. Biological Cybernetics 33: 237-247.
Freeman, W.J. (1987a). Techniques used in the search for the physiological basis of the EEG. In: Handbook of electroencephalography and clinical neurophysiology, vol. 3A, eds. A.S. Gevins and A. Remond. Amsterdam, Elsevier.
Freeman, W.J. (1987b). Simulation of chaotic EEG patterns with dynamic model of the olfactory system. Biological Cybernetics 56: 139-150.
Freeman, W.J. (1988). Strange attractors that govern mammalian brain dynamics shown by trajectories of electroencephalographic (EEG) potential. IEEE Transactions on Circuits and Systems. 35: 781-783.
Freeman, W.J. (1991). Predictions on neocortical dynamics derived from studies in paleocortex. In: Induced rhythms of the brain, eds. E. Basar & T.H. Bullock. Cambridge MA, Birkhaeuser Boston Inc.
Freeman, W.J. & Jakubith, S. (1993). Bifurcation analysis of continuous time dynamics of oscillatory neural networks. In: Brain Theory, ed. A. Aertson. Berlin, Springer-Verlag.
Freeman, W.J. & Skarda, C.A. (1985). Spatial EEG patterns, nonlinear dynamics and perception: the neo-Sherringtonian view. Brain Research Reviews 10: 147-175.
Gevins, A.S., Schaffer, R.E., Doyle, J.C., Cuttilo, B.A., Tannehill, R.S., Bressler, S.L. (1983). Shadows of thought: shifting lateralisation of human brain electrical patterns during a brief visuomotor task. Science 220: 97-99.
Gray, C.M., Koenig, P., Engel, K.A. & Singer, W. (1989). Oscillatory responses in cat visual cortex exhibit intercolumnar synchronisation which reflects global stimulus properties. Nature 338: 334-337.
Griniasty, M., Tsodyks, M.V. & Amit, D.J. (1993). Conversion of temporal correlations between stimuli to spatial correlations between attractors. Neural Computation 5: 1-17
Hopfield, J.J. (1982). Neural networks and physical systems with emergent collective computational abilities. Proceedings of National Academy of Sciences 79: 2558.
Hopfield, J.J. (1984). Neurones with graded response have collective computational properties like those of two state neurones. Proceedings of National Academy of Science 81: 3088-3092.
Ingber, L. & Nunez, P.L. (1990). Multiple scales of statistical physics of neocortex: application to electroencephalography. Math. Comput. Modelling 13: 83-95.
Hopfield, J.J. & Tank, D.W. (1986). Computing with neural circuits: a model. Science 233: 625-633.
Kaneko, K. (1990). Globally coupled chaos violates the law of large numbers. Physical Review Letters 65: 1391-1394.
Kaneko, K. (1992). Mean field fluctuation in network of chaotic elements. Physica D 55: 368-384.
Langton, C.G., Taylor, C., Farmer, J.D., & Rassmussen, S. (eds) (1992). Artificial Life II Santa Fe Institute Studies in the Sciences of Complexity, Proceedings vol 10. Redwood City, Addison-Wesley.
Liley, D.T.J. & Wright, J.J. (1994).
Intracortical connectivity of pyramidal and stellate cells:
estimates of synaptic densities and coupling symmetry.
Network: Computation in Neural Systems 5:175-189.
Little, W.A. & Shaw, G.L. (1975). A statistical theory of short and long term memory. Behavioural Biology 14: 115-133.
Lopes da Silva, F.H. & Storm van Leeuwen, W. (1978). The cortical alpha rhythm in dog: the depth and surface profile of phase. In: Architectonics of the Cerebral Cortex, eds. M.A.B. Brazier & H. Petsche. New York, Raven Press, pp.319-333.
Miyashita, Y. (1988). Neuronal correlate of visual associative long-term memory in the primate temporal cortex. Nature 335: 817-820.
Miyashita, Y. & Chang, H.S. (1988). Neural correlate of pictorial short-term memory in the primate temporal cortex. Nature 331: 68-70.
Nadal, J.P., Toulouse, G., Changeux, J.P. & Dehaene, S. (1986). Biophysics Letters 1(10): 535-542.
Nebenzahl, I. (1987). Recall of associated memories. Journal of Mathematical Biology 25: 511-519.
Nunez, P.L. ed. (1981). Electric fields of the brain. The neurophysics of EEG. New York, Oxford University Press.
Nunez, P.L. (1989). Generation of human EEG by a combination of long and short range neocortical interactions. Brain Topography 1: 199-215.
Nunez, P.L. & Srinivasan, R. (1993). Implications of recording strategy for estimates of neocortical dynamics with electroencephalography. Chaos 3: 257-266.
Nunez, P.L. (1994). Neocortical Dynamics and Human EEG Rhythms. New York, London, Oxford University Press.
Parisi, G. (1986a). A memory which forgets. Phys. A: Math. Gen.19: L617-L620.
Parisi, G. (1986b). Asymmetric neural networks and the process of learning. Phys. A: Math. Gen. 19: L675-L680.
Peretto, P. & Niez, J.J. (1986). Collective properties of neuronal networks. In: Disordered systems and biological organisation, eds. E. Bienenstock, F. Fogelman-Soulie, & G. Wiesbuch. Berlin, Springer-Verlag.
Picton, T.W. & Hillyard, S.A. (1988). Endogenous event related potentials. In: Picton, T.W. (ed) EEG Handbook. Amsterdam, Elsevier.
Ruelle, D. (1994). Physics Today 47: 24-30.
Sakai, K. & Miyashita, Y. (1991). Neural organisation for the long-term memory of paired associates. Nature 354: 152-155.
Schiff , J.S., Jerger, K., Duong, D.H., Chang, T., Spano, M.L., & Ditto, W.L. (1994). Controlling chaos in the brain. Nature 370: 615-620.
Sholl, D.A. (1953). Dendritic organization in the neurones of the visual and motor cortices of the cat. Journal of Anatomy 87: 387-407.
Silberstein, R.B. (1994). Neuromodulation of neocortical dynamics. In Nunez, P.L. Neocortical Dynamics and Human EEG Rhythms. New York, London, Oxford University Press.
Steriade, M., Gloor, P., Llinas, R.R., Lopes da Silva, F.H., & Mesulam, M.M. (1990). Basic mechanisms of cerebral rhythmic activities. Electroencephalography and clinical Neurophysiology 76: 481-508.
Stryker, M.P. (1989). Is grandmother an oscillation? Nature 338: 297-298.
Thatcher, R.W., Krause, P.J., & Hrybyk, M. (1986). Cortico-cortical associations and EEG coherence: a two-compartmental model. Electroenceph. clin. neurophysiol. 64: 123- 143.
Tovee, M.J. & Rolls, E.T. (1992). Oscillatory activity is not evident in the primate temporal visual cortex. Neuroreport 3: 369-372.
Tsuda, I. (1992) Dynamic link of memory-chaotic memory maps in non-equilibrium neural networks. 5: 313-326.Tsuda, I., Koerner, E. & Shimuzu, H. (1987). Memory dynamics in asynchronous neural networks. Progress in Theoretical Physics 78: 51-71.
Tsuda, I. (1994). Can stochastic renewal of maps be a model for cerebral cortex? Physica D (In press)
Uttley, A.M. (1956). The probability of neural connexions. Proceedings of the Royal Society B 142: 229-241.
Wilson, M.A. & Cowan, J.D. (1973). A mathematical theory of the functional dynamics of cortical and thalamic nervous tissue. Kybernetik 13: 55-80.
Wright, J.J. (1990).
Reticular activation and the dynamics of neuronal networks.
Biological Cybernetics 62: 289-298.
Wright, J.J., Kydd, R.R & Sergejew, A.A. (1990a).
Autoregression models of EEG.
Biological Cybernetics 62: 201-210.
Wright, J.J., Sergejew, A.A. & Stampfer, H.G. (1990b).
Inverse filter computation of the neural impulse giving
rise to the auditory evoked potential.
Brain Topography 2: 293- 302.
Wright, J.J. & Sergejew, A.A. (1991).
Radial coherence, wave velocity and damping of electrocortical waves.
Electroencephalography and clinical
Neurophysiology 79: 403- 412.
Wright, J.J. & Liley, D.T.J. (1994).
A millimetric-scale simulation of electrocortical wave
dynamics based on anatomical estimates of cortical synaptic density.
Network: Computation in Neural Systems 5: 191-202.
Wright,J.J., Sergejew,A.A., & Liley, D.T.J. (1994).
Computer simulation of electrocortical activity at
millimetric scale.
Electroencephalography and clinical Neurophysiology 90: 365-375.
Wright, J.J. & Liley, D.T.J. (1995).
Simulation of electrocortical waves.
Biological Cybernetics 72: 347-356.