Induction of Maternal Behavior in Rats: Effects
of
Pseudopregnancy Termination and Placenta-Smeared Pups
M. A. Steuer, A. C. Thompson, J. C. Doerr, M. Youakim,
and M. B. Kristal
State University of New York at Buffalo
Pseudopregnancy (PsP), a state hormonally similar to pregnancy (cessation of estrous cyclicity, elevated serum progesterone levels, diurnal and nocturnal prolactin surges), can be induced in the female rat by vaginocervical stimulation during estrus. Such stimulation enables prolactin surges to continue and the corpus luteum to be maintained in a functional state for approximately 14 days (Gunnet & Freeman, 1983; Pepe & Rothchild, 1974; Zarrow, Yochim, & McCarthy, 1964). Toward the end of PsP, as luteolysis develops, ovarian progesterone secretion declines, prolactin surges cease, ovarian estrogen secretion is enhanced, and normal estrous cycling resumes (Gunnet & Freeman, 1983; Hall & Robinson, 1978; Lau, Saksena, & Chang, 1979).
At the termination of PsP, several species exhibit some of the same "maternal" responses that they normally exhibit at the end of pregnancy. These responses have been reported in mice (Gandelman, McDermott, Kleinman, & Delianne, 1979; Noirot, Goyens, & Buhot, 1975), rabbits (Farooq, Denenberg, Ross, Sawin, & Zarrow, 1963; Hammond, cited in Klein, 1952, p. 88; Ross, Sawin, Zarrow, & Denenberg, 1963; Sawin, Denenberg, Ross, Hafter, & Zarrow, 1960; Tietz, 1933; Zarrow et al., 1961), cats (Tonkin, cited in Leyhausen, 1979, p. 89; McDonald, 1980), lions (Cooper, cited in Schneirla, Rosenblatt, & Tobach, 1963), and dogs (Brown, 1984; Mallo, 1971; Olson, Husted, Allen, & Nett, 1984; Smith & McDonald, 1974; Voith, 1980; Whitehead, 1966; Whitney, 1967).
Apparently only two studies have examined the effect of PsP termination on maternal responsiveness in laboratory rats. Terkel (1974) reported that normal (natural or spontaneous) PsP termination and pharmacological PsP termination induced by an injection of ergocornine induce maternal responsiveness in Sprague-Dawley rats within 48 hours of the end of PsP. Graber and Kristal (1977) found that the onset of maternal behavior in late-pseudopregnant nulliparous Long- Evans rats did not occur significantly sooner than in cycling nulliparous controls; however, when uterine distention was applied on Day 10 or 11 of PsP, maternal behavior appeared within 48 hours after the uterine-distention procedure. Graber and Kristal speculated that uterine distention had facilitated the onset of maternal behavior because it led to the release of prostaglandin-F2a (PGF2a ) which is known to cause luteolysis, a consequent reduction in progesterone, and therefore the termination of PsP (Hall & Robinson, 1978; Lau et al., 1979).
At term, virtually all parturient nonaquatic mammals are attracted to, and eventually ingest, the placenta, fetal membranes, and amniotic fluid associated with each neonate (for review, see Kristal, 1980). Kristal (1980) hypothesized that such attraction to placenta facilitated the onset of maternal behavior in laboratory rats by bringing the parturient female into immediate contact with each pup as it is born. Kristal, Whitney, and Peters (1981) then demonstrated, in rats, that placentophagic, nonpregnant, nulliparae retrieve and mother foster pups smeared with placenta much sooner than they do unsmeared pups or pups smeared with a cookie mash; thus, attraction of the adult to placenta on the foster young dramatically accelerates the onset of maternal responsiveness.
The present series of studies was undertaken to determine if the combination of PsP termination and presentation of placenta-smeared pups is a useful procedure with which to study the biobehavioral regulation of maternal behavior. We sought to determine which method of PsP termination in the Long-Evans rat reliably induces maternal behavior in the shortest amount of time, and to use that method to shed more light on some of the hormonal and experiential determinants of the onset of maternal behavior in the laboratory rat.
In Experiment 1, we examined the effects of various surgical PsP-termination
procedures on the induction of maternal behavior and determined whether
these effects were dependent upon the presence of ovarian estrogen. In
Experiment 2, we examined the effects of natural (spontaneous) PsP termination
on the onset of maternal behavior and determined whether this effect is
dependent upon estrogen. In Experiment 3, we investigated whether there
is a synergistic effect of PsP termination and placenta-induced attraction
to pups on the rapidity of the induction of maternal behavior.
General Method
Subjects
Subjects were 161 nulliparous female Long-Evans (hooded) rats, approximately 90-120 days old, and weighing approximately 250 g. All were kept on a 14:10 hr light/dark schedule, with lights on at 0700 hour (EST). Except where noted, all rats were housed individually in 24.5 x 18 x 18-cm galvanized wire mesh cages. Food (Agway Prolab Rat/Mouse/Hamster Formula 3000) and water were always available.
All subjects were born in our laboratory to parents purchased from Blue Spruce Farms (Altamont, NY). At approximately 60 days of age, all potential subjects were screened for regular ovarian cyclicity by daily inspection of vaginal smears; noncycling females were excluded from further study. All were tested for spontaneous placentophagia by the method described by Kristal and Graber (1976), and except where noted in Experiment 3, all were determined to be nonplacentophages.
All potential subjects then underwent a second pretest to screen out those that exhibit either spontaneous maternal behavior (approximately 4% in our Long-Evans virgins) or spontaneous infanticide of stimulus pups (approximately 10%). Following the method described by Kristal et al. (1981), 4 stimulus pups, 3-8 days of age, were placed in the cage of each rat. Rats that retrieved any of the pups during the 15-min exposure period were judged to be spontaneous retrievers; females that bit or began to eat any of the pups were judged to be spontaneous pup-killers/eaters. Both spontaneous retrievers and spontaneous pup-killers/eaters were excluded from further study.
A daily vaginal smear from each rat was obtained and analyzed at 1000 hour. Pseudopregnancy (PsP) was induced by mating the subjects with vasectomized adult males on the evening of proestrus. On the following morning at 1000 hour, the area beneath the male's cage was checked for the presence of fallen vaginal plugs; the female was also inspected for the presence of a vaginal plug, and then returned to her home cage. Successful induction of PsP was indicated by the presence of a vaginal plug on the morning following the sterile mating (Day I of PsP). Maintenance of PsP was confirmed by daily vaginal smear records; a rat was considered to be pseudopregnant only if she remained in diestrus for the 12 days following the sterile mating.
Maternal-Behavior Testing
All maternal-behavior tests were conducted in clear, wire-topped plastic cages. 21 x 42 x 20 cm, containing approximately 3 cm of coarse sawdust. Food and water were available ad lib. Stimulus pups used for maternal-behavior testing were provided by a bank of Long- Evans donor mothers.
Each rat was tested for maternal behavior twice daily. At 1000 hour, each rat was presented with four fresh, 3- to 8-day-old stimulus pups, and was observed for the next 15 min. The second test occurred at 2200 hour; the same pups were scattered about the cage. A rat was considered to be maternal only if she had retrieved, licked, and crouched over all four stimulus pups during each of two consecutive tests. The number of hours or days to reach criterion for maternal behavior on the first of the two criterion tests is referred to as the "maternal-behavior latency." The stimulus pups remained with the rat until replaced by four fresh foster pups the next day. On the first day of continuous exposure to pups, several spot checks for the rapid onset of maternal behavior were made throughout the day.
The twice-daily maternal-behavior tests were continued until each rat behaved maternally or until 7 days of testing had elapsed. Any rat that had not behaved maternally by the end of the seventh day was assigned a latency of 8 days.
Method
Subjects
Sixty-four maternally naive virgin females were made pseudopregnant. On Day 12 of PsP, they were divided evenly into four groups, and each rat received one of the following surgical procedures: ovariectomy (0), combined ovariectomy and hysterectomy (OH), hysterectomy control (H), or sham surgery (SS). Half of the females in each surgical group (n = 8) then received a subcutaneous injection of 50 m g/kg estradiol benzoate (EB), and the other half received olive oil vehicle (OIL). (The 50 m g/kg EB dose was used because it is in the middle of the range of effective doses for estrogen-induced facilitation of maternal behavior [Fahrbach & Pfaff, 1982; Rosenblatt et al., 1979].) Then each rat was placed in a clear plastic test cage and allowed to recover. Vaginal smears were obtained daily from each rat.
Surgical Procedures
All operations were performed between 1000 and 1130 hour on Day 12 of PsP. Rats were anesthetized by injection (0.65 ml/kg, ip) of a 4:1 mixture of ketamine hydrochloride and xylazine hydrochloride (Ketaset, 100 mg/ml, Bristol Laboratories; Rompun, 20 mg/mI, Cutter Laboratories). Ovariectomy was performed through bilateral 1.0- to 1.5-cm lateral incisions through the skin and abdominal wall; the ovaries were pulled through these openings, ligated, and removed. Hysterectomy was performed by making a 3-cm ventral midline incision through the skin and abdominal wall, approximately 3 cm anterior to the vaginal orifice; the uterus and cervix were pulled through this opening, ligated and removed, leaving the ovaries and vagina in the rat. The surgical procedure for combined ovariectomy + hysterectomy was identical to that used for hysterectomy except that both the ovaries and the uterus were ligated and removed. Sham surgery was performed through a 3-cm midventral incision; the abdominal cavity was opened, but neither the ovaries nor the uterus were touched. In all rats, the abdominal wall was closed with 00 silk suture, and the skin with 000 silk suture.
Procedure
Behavioral testing, as described in the General Method, began 48 hr after surgery on Day 14 of PsP.
Results and Discussion
The median duration of PsP for intact Long-Evans rats in our laboratory is 14 days (see Experiment 3B). Among ovariectomized rats, the median duration of PsP for rats that received exogenous estrogen was not different from normal (0 + EB, 14 days; OH + EB, 14 days); the median duration of PsP for ovariectomized rats that did not receive exogenous estrogen (0 + OIL; OH + OIL) could not be determined because these rats never exhibited vaginal cornification. Among rats with intact ovaries, the median duration of PsP was normal in surgical groups that received exogenous estrogen (H + EB, 14.5 days; SS + EB, 16.5 days) and was longer than normal in surgical groups that did not receive exogenous estrogen (H + OIL, 17.5 days; SS + OIL, 16.0 days). It should be noted that hysterectomy alone prolongs rather than terminates PsP.
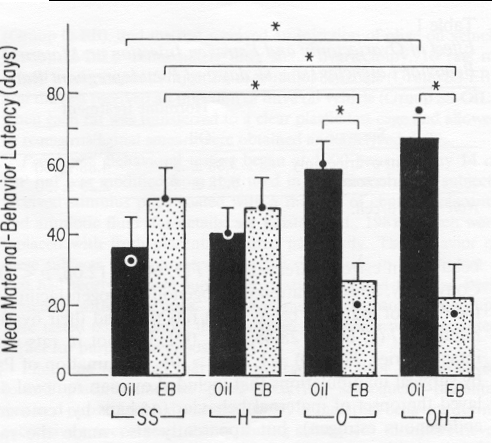
Figure 1. Mean number of days (± SEM) to the onset of maternal behavior in pseudopregnant rats that underwent either sham surgery (SS), hysterectomy (H), ovariectomy (O), or ovariectomy + hysterectomy (OH), and were injected subcutaneously with oil (Oil) or estradiol benzoate (EB). (8/group. · = median.) * p < .05.
Figure 1 depicts the results of the first study. The overall analysis
of variance (ANOVA) revealed a significant Surgery x
Hormone interaction. F(3,56) = 7.21, p <
.01. Subsequent tests involved one-way ANOVAs with appropriately adjusted
mean square errors and Newman-Keuls pairwise comparisons (ps <
.05).
Tests of the Surgery x Drug interaction yielded significant effects of surgery in oil-treated rats, F(3,56) = 3.48, p < .025, as well as in estrogen-treated rats, F(3,56) = 3.82, p < .025. In oil-treated rats, those in the OH group had significantly longer maternal-behavior latencies than those in the SS group; the O group and H group latencies were not different from those of the SS group. In estrogen-treated rats, the OH group and O group latencies were significantly shorter than those of the SS group; the H group had latencies that were not different from those of the SS group.
Tests of the Surgery x Hormone interaction showed that in comparison to oil injection, estrogen injection hastened the onset of maternal behavior in the OH group, F(1,56) = 17.27, p < .001, and the O group, F(1,56) = 10.14, p < .001, but did not do so in the H group, F(1,56) < 1.00, or the SS group, F(1,56) = 1.43, p > .05.
In a separate analysis aimed at determining the extent to which the
presence or absence of ovaries affects maternal behavior in late-PsP rats,
we reanalyzed the data in terms of only two surgical groups: rats with
intact ovaries (H and SS groups) and rats without ovaries (O and OH groups).
The results of this analysis are depicted in Table 1. The overall analysis
revealed a significant Surgery x Hormone interaction,
F(1,60) = 22.13, p < .01. Removal of the ovaries
inhibited the onset of maternal behavior in oil-treated rats, F(1,60)
= 10.14, p < .01, but facilitated the onset of maternal behavior
in estrogen-treated rats, F(1,60) = 12.06, p < .01.
Conversely, administration of exogenous estrogen facilitated the onset
of maternal behavior in rats that had their ovaries removed, F(1,60)
= 28.45, p < .001, but not in rats with intact ovaries,
F(1,60) = 1.75, p > .05. Termination of PsP by surgical
manipulations that included ovarian removal delayed the onset of maternal
behavior (perhaps by removing endogenous estrogen), but apparently also
made the rats hypersensitive to the effects of exogenous estrogen.
Table 1
Effect of Ovariectomy and Estrogen Injection on Maternal
Behavior Latencies (M ± SE
days) of Pseudopregnant Rats
|
|
||
|
|
|
|
| Sham ovariectomy |
|
|
| Ovariectomy |
|
|
| Note: n = 16/cell | ||
Experiment 2: Natural PsP Termination and Maternal Behavior
Although the results of Graber and Kristal (1977) and Terkel (1974) showed that experimental termination of PsP facilitates the onset of maternal behavior, those studies offer different information on the effect of natural termination of PsP on the onset of maternal behavior: Terkel found a facilitation of the onset of maternal behavior after natural PsP termination, but Graber and Kristal did not. Graber and Kristal initiated maternal-behavior testing on Day 10 of PsP, which is approximately 4 days before PsP terminates naturally (Gunnet & Freeman, 1983; Kristal & Steuer, 1984). It is possible that if behavioral testing had begun closer to the termination of PsP, Graber and Kristal would have seen maternal behavior sooner. It is also possible that because natural termination of PsP produces a more "physiological" sequence of changes in ovarian steroid levels than does surgical termination of PsP, such natural termination of PsP may facilitate the onset of maternal behavior as well as, or even more effectively than, any invasive PsP-termination procedure. Therefore, in the present study, we initiated maternal-behavior testing closer to the end of PsP (on Day 14) in PsP subjects and simply allowed PsP to terminate naturally. In addition, we tested whether the concomitant administration of estrogen helps hasten the onset of maternal behavior in unoperated rats, because all the rats in Experiment 1 had undergone anesthetization and surgery.
Method
Subjects
Twenty-eight maternally naive virgin female rats were used. Sixteen were made pseudopregnant (PsP group); 12 remained unmated and served as cycling controls (CYC group). On Day 12 of PsP, 8 PsP rats and 6 CYC rats received an injection of 50 m g/kg estradiol benzoate; the remaining rats in both groups received an injection of olive oil vehicle. All injections were administered between 1000 hour and 1130 hour. After injection, each rat was moved to a clear plastic test cage. A vaginal smear from each rat was obtained and analyzed daily.
Procedure
Behavioral testing began 48 hours after the injection on Day 14 of PsP.
Results and Discussion
Figure 2 depicts the results of the second study. The overall ANOVA revealed only a significant main effect of reproductive state, F(1,24) = 45.24, p < .01; PsP rats exhibited maternal behavior sooner than CYC rats. Treatment with exogenous estrogen did not affect the onset of maternal behavior in either PsP or CYC rats, F(1,24) < 1.0.
Vaginal smear records of the six PsP nulliparae that were not treated with exogenous estrogen suggested that the resumption of estrous cyclicity after PsP termination may be associated with the onset of maternal responsiveness, because all 6 PsP rats manifested an estrus vaginal smear within 24 hours of the onset of maternal behavior. (Experiment 3B examines this relation more systematically.) We did not detect the presence of a similar relation in estrogen-treated PsP nulliparae, because estrous cyclicity was disrupted in 5 of 6 rats that received estrogen (4 remained in diestrus, 1 exhibited 3 days of estrus followed by constant diestrus).
Figure 2. Mean number of days (± SEM) to the onset of maternal behavior in rats showing estrous cyclicity (CYC) or undergoing natural termination of pseudopregnancy (PsP) and that were injected subcutaneously with oil (Oil) or estradiol benzoate (EB). (Number in parentheses = n; · = median. *p < .05.)
Experiment 3: Effect of Placenta-Smeared Pups on
Maternal Behavior After PsP Termination
In Experiment 1, we showed that ovariectomy + estrogen injection and ovariectomy + hysterectomy + estrogen injection facilitate the onset of maternal behavior in PsP rats, presumably by combining a rapid reduction of progesterone with the increase in estrogen. In Experiment 2, we showed that natural termination of PsP also facilitates the onset of maternal behavior. Taken together, these two experiments suggest that PsP termination is as effective in facilitating the onset of maternal behavior as pregnancy termination is re ported to be. After PsP termination or pregnancy termination, however, rats require several hours of exposure to standard stimulus pups (from 6 to 48 hr) before they exhibit maternal behavior (Fahrbach & Pfaff, 1982; Graber & Kristal, 1977; Rosenblatt et al., 1979; Stern, 1983; Terkel, 1974).
The duration of exposure to pups necessary before non-pregnant rats
display maternal behavior can be reduced by manipulations that alter the
stimulus quality of foster pups or by manipulations that presumably alter
the way in which those foster pups are perceived (Fleming & Rosenblatt,
1 974a, 1974b; Fleming, Vaccarino, Tambosso, & Chee, 1979; Kristal
et al., 1981; Mayer & Rosenblatt, 1980; Stern, 1983; Terkel & Rosenblatt,
1971). At parturition, rats have undergone dramatic changes in blood levels
of estrogen and progesterone; they are now also placentophagic, and for
the first time are exposed to and immediately attracted to the placenta,
fetal membranes, and amniotic fluid associated with neonates. Kristal et
al. (1981) showed that this attraction to the afterbirth hastens the onset
of maternal behavior in cycling nulliparae. Presentation of placenta-covered
stimulus pups to PsP-terminated rats would simulate the environmental cues
as well as the hormonal milieu present at parturition, Therefore, in Experiment
3, we tested whether the attraction to placenta, in combination with the
hormonal changes resulting from PsP termination + estrogen injection, induces
maternal behavior more rapidly than does PsP termination + estrogen injection
alone. We tested for this combined effect using nulliparae in which PsP
had been terminated by ovariectomy (Experiment 3A), and in which PsP had
terminated naturally (Experiment 3B).
Experiment 3A: Surgical PsP Termination and
Maternal Behavior Toward Placenta-Smeared Pups
Method
Subjects. Forty-one maternally naive virgin females were used; all had been determined to be placentophages, using the method described by Kristal and Graber (1976), or had been induced to eat placenta after 4-8 exposures to donor placenta coupled with food deprivation; once a rat has eaten placenta in this manner, it will reliably do so when subsequently presented with donor placenta (Kristal, 1980; Kristal & Graber, 1976). All were made pseudopreg nant. On day 12 of PsP, 21 rats underwent ovariectomy and 20 underwent sham-ovariectomy. Immediately after ovariectomy, 13 rats received a subcutaneous injection of 50 m g/kg estradiol benzoate (Group O-EB), and the rest received an injection of olive oil vehicle (Group O-OIL). Immediately after sham ovariectomy, 10 rats received a subcutaneous injection of estradiol benzoate (Group SS-EB), and the rest received an injection of olive oil vehicle (Group SS-OIL). Then each rat was transferred to a clear plastic test cage and allowed to recover. Vaginal smears were obtained and analyzed daily.
Procedure. Behavioral testing began at 1000 hour on Day 14 of PsP but was modified from that used in Experiment 1: all subjects received stimulus pups coated with a mixture of chopped placenta and amniotic fluid (for details, see Kristal et al., 1981), which were replaced with fresh placenta-smeared pups daily. The behavior of these subjects was compared with that of the rats in Experiment I that had received sham surgery or ovariectomy and that had been presented with clean pups. The clean-pup and smeared-pup groups were tested concurrently; the clean-pup group data were presented earlier for purposes of exposition.
Pilot data indicated that once maternal behavior was displayed toward placenta-smeared pups, receiving subsequent placenta-smeared pups interfered with the display of maternal behavior during successive retrieval tests; that is, during subsequent 1 5-min retrieval tests, subjects were more interested in licking placenta and fluids from the pups than in retrieving them. Therefore, the subjects' behavior toward the pups was monitored for 2 hr after each test. At the end of 2 hr, any rat exhibiting maternal behavior (crouching over all four pups) was given another 15-min test using clean pups. (Taken together, these procedures simulate conditions at parturition, when mother rats are exposed to amniotic-fluid-covered and placenta-covered pups, they lick them clean and then behave maternally toward the cleaned pups.)
If the rat began to bite or eat the pups during the 2-hr maternal-behavior test, the pups were removed and she was given new pups at the next maternal-behavior test (12 hr later). Data from such infanticidal rats were analyzed separately from those of noninfanticidal rats because the former group did not receive continuous exposure to pups, whereas the latter group did. Unless otherwise stated, the results reported are only from noninfanticidal rats.
Results and Discussion
Figure 3 depicts the results of Experiment 3A. In the following analyses, we used data from ovariectomized and sham ovariectomized rats that were continuously exposed to placenta-smeared pups (this experiment) and data from ovariectomized and sham-ovariectomized rats that were continuously exposed to clean pups (Experiment 1). (The only difference between the two experiments is the type of pups that were proffered: clean or placenta-smeared.)
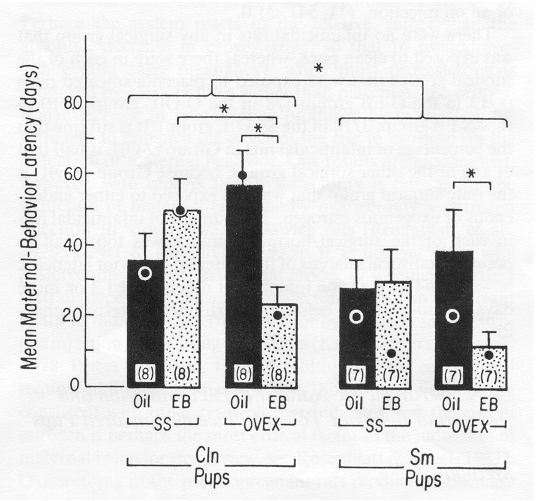
The overall ANOVA revealed a significant Surgery x Hormone interaction, F(1,54) = 9.51, p < .01. The main effect of stimulus-pup type, F(1, 54) = 6.24, p < .025, indicated that rats exposed to placenta-smeared pups became maternal significantly sooner than did rats exposed to clean pups.
The Surgery x Hormone interaction was probed for each pup type (clean pups, placenta-smeared pups) using one-way ANOVAs with adjusted mean square errors. Among rats exposed to clean pups, there was a significant effect of surgery in those that received estrogen, F(1,54) = 5.06, p < .05, but not in those that received oil, F(1,54) = 3.16, p > .10. Therefore, as in Experiment 1, ovariectomy resulted in significantly shorter maternal-behavior latencies than sham surgery only in estrogen-treated rats.
Figure 3. Mean number of days (± SEM) to the onset of maternal behavior in pseudopregnant rats that underwent either sham surgery (SS) or ovariectomy (OVEX), were injected subcutaneously with either oil (Oil) or estradiol benzoate (EB), and were presented with clean pups (CIn) or pups smeared with placenta and amniotic fluid (Sm). (Number in parentheses = n; · = median. *p < .05.)
In contrast to rats exposed to clean pups, those exposed to placenta-smeared pups showed no effect of surgery whether they received an estrogen injection, F(1,54) = 1.84, p > .10, or an oil injection, F(1, 54) < 1.0.
There were no infanticidal rats in any surgical group that was exposed
to clean pups, whereas there were in each of the surgical groups that were
exposed to placenta-smeared pups (4/13 in the O-EB group; 1/8 in the O-OIL
group; 3/10 in the SS-EB group; 3/10 in the SS-OIL group). It is striking
that the percentage of infanticidal rats in Group O-OIL is half that of
any of the other surgical groups, because Group O-OIL is the only surgical
group that was not exposed to either endogenous or exogenous estrogen.
The number of infanticidal rats in each of the surgical/hormone groups
was too small to permit a statistical analysis of the maternal-behavior
latencies; however, 64% of all the infanticidal rats required 3 or more
days of pup-exposure before they exhibited maternal behavior.
Experiment 3B: Natural PsP Termination and
Maternal Behavior Toward Placenta-Smeared Pups
Method
Subjects. Twenty-eight maternally naive virgin placentophagic females were used. Fourteen of the subjects were made pseudopregnant (Groups PsP-T), and 14 remained unmated and served as cycling controls (Group CYC). On Day 10 of PsP, each subject was transferred to a clear plastic test cage containing approximately 3 cm of coarse sawdust. A daily vaginal smear from each rat was obtained and analyzed at 1000 hour and 2200 hour.
Procedure. The first proestrus or cornified vaginal smear was taken as evidence of the termination of PsP. Behavioral testing began as soon as PsP had ended in each rat. During behavioral testing, 8 of the PsP-T rats received placenta-smeared stimulus pups, and the rest received clean stimulus pups; eight of the CYC rats received placenta-smeared stimulus pups, and the rest received clean stimulus pups. Pups were replaced with fresh pups (either placenta-smeared or clean) at 12-hr intervals. Unless otherwise stated, the results reported are only from noninfanticidal rats.
Results and Discussion
The median duration of PsP for all PsP-T rats was 14.0 days. There was no significant difference between the length of PsP in rats exposed to placenta-smeared pups (14.5 days) and that of rats exposed to clean pups (14 days).
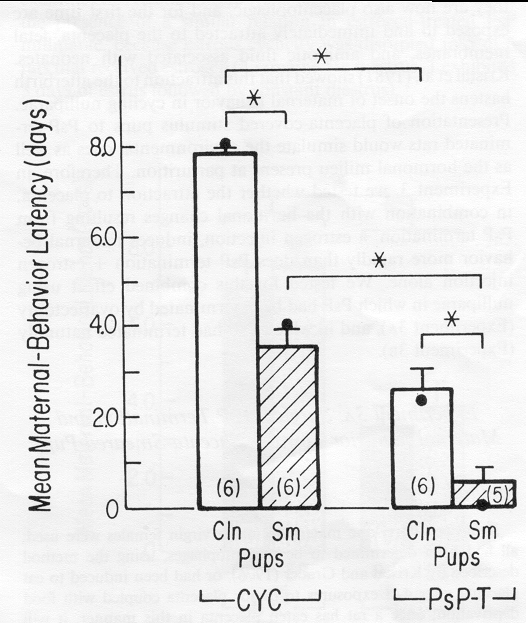
Figure 4 depicts the results of Experiment 3B. There was a significant interaction between reproductive condition and pup type, F(l, 19) = 5.84, p < .05.
Tests of the interaction revealed that exposure to placenta- smeared pups produced maternal behavior significantly sooner than exposure to clean pups, both in PsP-T rats, F(1,19) = 8.86, p < .01, and in CYC rats, F(1,19) = 44.04, p < .01. In fact, the median latency for PsP-T rats to exhibit maternal behavior toward placenta-smeared pups was 2 hr whereas that toward clean pups was 54 hr. The median latency for CYC rats to exhibit maternal behavior toward placenta- smeared pups was 96 hr whereas that toward clean pups was 192 hr.
Figure 4. Mean number of days (± SEM) to the onset of maternal behavior in rats showing estrous cyclicity (CYC) or undergoing natural termination of pseudopregnancy (PsP-T) that were presented with either clean (CIn) pups or pups smeared with placenta and amniotic fluid (Sm). (Number in parentheses = n; · = median. *p <.05.)
PsP-T rats became maternal significantly sooner than did CYC rats, whether they were exposed to placenta-smeared pups, F(1,19) = 21.70, p < .01, or to clean pups, F(1,19) = 69.10, p< .01.
PsP-T rats presented with clean pups after PsP termination became maternal 17 days after the initiation of PsP (15 days of PsP + 2 days of pup exposure after PsP had terminated). Because PsP lasts approximately 15 days (shown in this experiment), it is now apparent that PsP rats in Experiment 2 were first presented with clean pups 1 day prior to PsP termination -- that is, on Day 14 of PsP; nevertheless, these rats also became maternal 17 days after the initiation of PsP (14 days of PsP + 1 day of pup exposure on the last day of PsP + 2 days of pup exposure after PsP had terminated). Therefore, once PsP had terminated, the rats in Experiment 3B and in Experiment 2 became maternal at approximately the same time -- after 2 days of exposure to clean pups. This suggests that in Experiment 2, exposing rats to clean pups during the last day of PsP may have been superfluous, that is, that exposing rats to clean pups for 2 days may facilitate the onset of maternal behavior only after those rats have under gone PsP termination. However, it is important to note that exposing rats to placenta-smeared pups, rather than to clean pups, reduces the duration of pup exposure necessary between PsP termination and the onset of maternal behavior from 2 days to 2 hours.
As in Experiment 3A, there were no rats among either PsP-T rats or CYC rats that were infanticidal when exposed to clean pups, but there were some PsP-T rats and CYC rats that were infanticidal when exposed to placenta-smeared pups (3/8 in the PsP-T group, 2/8 in the CYC group). Again, the incidence of infanticide was too low to permit a statistical analysis of the maternal-behavior latencies. However, all 5 of the infanticidal rats required 2.5 or more days of pup exposure before they exhibited maternal behavior.
General Discussion
The present series of studies was designed to investigate the effects of pseudopregnancy termination and exposure to placenta-smeared pups on the induction rate of maternal behavior in maternally naive Long-Evans rats. In Experiment 1 we demonstrated that (a) ovariectomy and an estrogen injection on Day 12 of pseudopregnancy, with or without a concomitant hysterectomy, hastened the onset of maternal behavior. Interestingly, (b) ovariectomy at that time, without an estrogen injection (with or without a concomitant hysterectomy), retarded the onset of maternal behavior, and (c) estrogen injection without an ovariectomy (with or without a concomitant hysterectomy) had no effect on maternal-behavior latency. Finally, (d) hysterectomy, whether accompanied by an estrogen or oil injection, had no effect on maternal-behavior latency.
In Experiment 2 we showed that (a) natural, rather than surgically induced, termination of pseudopregnancy is also associated with a rapid onset of maternal behavior. When maternal-behavior testing was begun on Day 14 of pseudopregnancy, the pseudopregnant rats required half as much exposure to pups to begin mothering them as did the control rats. (b) In naturally terminating pseudopregnant rats, in contrast to the ovariectomized rats of Experiment 1, estrogen injection had no effect on maternal-behavior latency.
In Experiment 3, each of the two effective procedures, ovariectomy-induced termination of pseudopregnancy (in conjunction with an estrogen injection) and natural termination of pseudopregnancy, was combined with the presentation of placenta-smeared stimulus pups. Previous research has shown that placenta-smeared pups are mothered sooner than clean pups by virgin rats attracted to placenta (Kristal et al., 1981). We found that (a) placenta-smeared pups produced shorter maternal-behavior latencies than did clean pups in all groups, and (b) when combined with the presentation of placenta-smeared pups, natural termination of pseudopregnancy produced shorter latencies than did the surgical-termination/estrogen-injection procedure. The median latency for the fastest group to become maternal was 2 hr.
The two effective PsP-termination conditions, ovariectomy + estrogen injection and natural termination, apparently re produce the hormonal changes of a terminating pregnancy sufficiently to induce the onset of maternal behavior after only a brief exposure to pups. The onset of maternal behavior after pseudopregnancy termination seems independent of uterine factors; hysterectomy did not influence the pseudo-pregnancy-termination effect found in Experiment 1, whereas hysterectomy (more probably removal of the progesterone secreting placenta) is a sine qua non for the pregnancy-termination effect (for review, see Rosenblatt et al., 1979). Perhaps the system reacts to the relative, rather than the absolute, decrease in progesterone, so that in pseudopregnancy (and early pregnancy), when most progesterone is produced by corpora lutea rather than placentas, the proportional decrease in progesterone produced by ovariectomy is sufficient to enhance the effect of the rise in estrogen on the induction of maternal behavior. Our results are inconsistent, however, with reports that hysterectomy is critical for the induction of maternal behavior by ovariectomy + estrogen injection in virgin Sprague-Dawley rats (Rosenblatt et al., 1979; Siegel & Rosenblatt, 1975). It is not yet clear why hysterectomy is necessary in that model, or whether the effect is strain dependent, because hysterectomy is not an important factor in inducing maternal behavior by ovariectomy + estrogen injection in our Long-Evans rats (Kristal, Peters, & Steuer, 1978; Peters & Kristal, 1980). Resolution of the conflict will require additional studies. Clearly, though, our findings are consistent with other research that indicates that the rise in estrogen is perhaps the most critical factor in the induction of maternal behavior (for review, see Rosenblatt & Siegel, 1981). Ovariectomy in the pseudopregnant rats produced a decrease in endogenous estrogen as well as progesterone. If estrogen was not replaced, the induction of maternal behavior was hindered. Replacement with exogenous estrogen (during the period of a rapidly diminishing progesterone level) produced a significant shortening of the maternal-behavior latency. It should be noted that simply raising estrogen levels by administering exogenous estrogen to pseudopregnant rats with ovaries did not have any effect on the baseline maternal-behavior latency.
Presentation of placenta-smeared pups to placentophagic rats at the
time that pseudopregnancy is terminating naturally apparently produces
a constellation of internal (hormonal) and external stimuli that approximates
those present during delivery, thereby generating a very rapid onset of
maternal behavior. This procedure should prove to be extremely useful in
research into the psychobiological determinants of maternal behavior because
(a) it produces maternal-behavior latencies as short as the shortest produced
by other procedures currently in use, and (b) it requires neither surgical
intervention nor the administration of exogenous hormones.
This research was supported in part by funds made available
to M. B. Kristal from BRSG S07 RR 07066, awarded to SUNY-Buffalo by the
Biomedical Research Support Grant Program, Division of Research Resources,
National Institutes of Health.
We thank Seymour Axelrod for his editorial advice.
Correspondence concerning this article should be addressed
to M. B. Kristal, Department of Psychology, SUNY at Buffalo, Buffalo, New
York 14260.
References
Bridges, R. S. (1984). A quantitative analysis of the roles of dosage, sequence, and duration of estradiol and progesterone exposure on the regulation of maternal behavior in the rat. Endocrinology, 114, 930-940.
Bridges, R. S., DiBiase, R., Loundes, D. D., & Doherty, P. C. (1985). Prolactin stimulation of maternal behavior in female rats. Science, 227, 782-784.
Brown, J. M. (1984). Efficacy and dosage titration study of mibolerone for treatment of pseudopregnancy in the bitch. Journal of the American Veterinary Medicine Association, 184, 1467-1468.
Farooq, A., Denenberg, V. H., Ross, S., Sawin, P. B., & Zarrow, M. X. (1963). Maternal behavior in the rabbit: Endocrine factors involved in hair loosening. American Journal of Physiology, 204, 271-274.
Fahrbach, S. F., & Pfaff, D. W. (1982). Hormonal and neural mechanisms underlying maternal behavior in the rat. In D. W. Pfaff (Ed.), The physiological mechanisms of motivation (pp. 253-285). New York: Springer-Verlag.
Fleming, A. S., & Rosenblatt, J. S. (1974a). Olfactory regulation of maternal behavior in rats: I. Effects of olfactory bulb removal in experienced and inexperienced lactating and cycling females. Journal of Comparative and Physiological Psychology, 86, 221-232.
Fleming, A. S., & Rosenblatt, J. S. (1974b). Olfactory regulation of maternal behavior in rats: II. Effects of peripherally induced anosmia and lesions of the lateral olfactory tract in pup-induced virgins. Journal of Comparative and Physiological Psychology, 86, 233-246.
Fleming, A., Vaccarino, F., Tambosso, L., & Chee, P. (1979). Vomeronasal and olfactory system modulation of maternal behavior in the rat. Science, 203, 372-374.
Gandelman, R., McDermott, N. J., Kieinman, M., & DeJianne, D. (1979). Maternal nest building by pseudopregnant mice. Journal of Reproduction and Fertility, 56, 697-699.
Graber, G. C., & Kristal, M. B. (1977). Uterine distention facilitates the onset of maternal behavior in pseudopregnant but not in cycling rats. Physiology and Behavior, 19, 133-137.
Gunnet; J. W., & Freeman, M. E. (1983). The mating-induced release of prolactin: An unique neuroendocrine response. Endocrine Reviews, 4, 44-61.
Hall, A. K., & Robinson, J. (1978). Progesterone, 20-a -dihydroprogesterone and PGF: Interrelated hormonal changes during pseudopregnancy in the rat. Prostaglandins, 15, 1013-1025.
Klein, M. (1952). Uterine distension, ovarian hormones and maternal behaviour in rodents. CIBA Foundation Colloquia on Endocrinology, 3, 84-88.
Kristal, M. B. (1980). Placentophagia: A biobehavioral enigma (or De gustibus non disputandum est). Neuroscience and Biobehavioral Reviews, 4, 141-150.
Kristal, M. B., & Graber, G. C. (1976). Placentophagia in nonpregnant rats: Influence of estrous cycle stage and birthplace. Physiology & Behavior, 17, 599-605.
Kristal, M. B., Peters, L. C., & Steuer, M. A. (1978). Estrogen and the initiation of placentophagia. Paper presented at the Conference on Reproductive Behavior, Madison, WS.
Kristal, M. B., & Steuer, M. A. (1984). [The duration of pseudopregnancy in Long-Evans rats]. Unpublished raw data.
Kristal, M. B., Whitney, J. F., & Peters, L. C. (1981). Placenta on pups' skin accelerates the onset of maternal behaviour in non-pregnant rats. Animal Behaviour, 29, 81-85.
Lau, I. F., Saksena, S. K., & Chang, M. C. (1979). Serum concentrations of progestins, estrogens, testosterone and gonadotropins in pseudopregnant rats with special reference to the effect of prostaglandin F2a,. Biology of Reproduction, 20, 575-580.
Leyhausen, P. (1979). Cat behavior: The predatory and social behavior of domestic and wild cats. New York: Garland STPM Press.
Mallo, G. L. (1971). False pregnancy. In R. W. Kirk (Ed.), Current veterinary therapy: Small animal practice: 1971 (Vol. 4, pp. 756-757). Philadelphia: W. B. Saunders.
Mayer, A. D., & Rosenblatt, J. 5. (1980). Hormonal interaction with stimulus and situational factors in the initiation of maternal behavior in nonpregnant rats. Journal of Comparative and Physiological Psychology, 94, 1040-1059.
McDonald, L. E. (1980). Veterinary endocrinology and reproduction (3rd ed.). Philadelphia: Lea and Febiger.
Noirot, E., Goyens, J., & Buhot, M. -C. (1975). Aggressive behavior of pregnant mice toward males. Hormones and Behavior, 6, 9-17.
Olson, P. N., Husted, P. W., Allen, T. A., & Nett, T. M. (1984). Reproductive endocrinology and physiology of the bitch and queen. Veterinary Clinics of North America: Small Animal Practice, 14, 927-946.
Pepe, G. J., & Rothchild, I. (1974). A comparative study of serum progesterone levels in pregnancy and in various types of pseudo-pregnancy in the rat. Endocrinology, 95, 275-279.
Peters, L. C., & Kristal, M. B. (1980). [The effect of hysterectomy on maternal behavior of Long-Evans rats]. Unpublished raw data.
Rosenblatt, I. S., & Siegel, H. I. (1981). Factors governing the onset and maintenance of maternal behavior among nonprimate mammals: The role of hormonal and nonhormonal factors. In D. J. Gubernick & P. H. Klopfer (Eds.), Parental care in mammals (pp. 14-76). New York: Plenum.
Rosenblatt, J. S., Siegel, H. I., & Mayer, A. D. (1979). Progress in the study of maternal behavior in the rat: Hormonal, nonhormonal, sensory, and developmental aspects. In J. S. Rosenblatt, R. A. Hinde, E. Shaw, & C. Beer (Eds.), Advances in the study of behavior (Vol. 10, pp. 225-311). New York: Academic.
Ross, S., Sawin, P. B., Zarrow, M. X., & Denenberg, V. H. (1963). Maternal behavior in the rabbit. In H. L. Rheingold (Ed.), Maternal behavior in mammals (pp. 94-121). New York: Wiley.
Sawin, P. B., Denenberg, V. H., Ross, S., Hafter, E., & Zarrow, M. X. (1960). Maternal behavior in the rabbit: Hair loosening during gestation. American Journal of Physiology, 198, 1099-1102.
Schneirla, T. C., Rosenblatt, J. S., & Tobach, E. (1963). Maternal behavior in the cat. In H. L. Rheingold (Ed.), Maternal behavior in mammals (pp. 122-168). New York: Wiley.
Siegel, H. I., & Rosenblatt, J. S. (1975). Estrogen-induced maternal behavior in hysterectomized-ovariectomized virgin rats. Physiology and Behavior, 14, 465-471.
Smith, M. S., & McDonald, L. E. (1974). Serum levels of luteinizing hormone and progesterone during the estrous cycle, pseudopregnancy and pregnancy in the dog. Endocrinology, 94, 404-412.
Stern, J. M. (1983). Maternal behavior priming in virgin and caesarean-delivered Long-Evans rats: Effects of brief contact or continuous exteroceptive pup stimulation. Physiology and Behavior,31, 757-764.
Terkel, J. (1974). Maternal behavior upon experimental and normal termination of pseudopregnancy in the rat. Paper presented at the 14th International Ethological Conference, Parma, Italy.
Terkel, J., & Rosenblatt, J. 5. (1971). Aspects of nonhormonal maternal behavior in the rat. Hormones and Behavior, 2, 161-171.
Tietz, E. B. (1933). The humoral excitation of the nesting instincts in rabbits. Science, 78, 316.
Voith, V. L. (1980). Functional significance of pseudocyesis. Modern Veterinary Practice, 61, 75-77.
Whitehead, J. E. (1966). Pseudopregnancy. In R. W. Kirk (Ed.), Current veterinary therapy: Small animal practice: 1966-1967 (Vol. 1, pp. 423-425). Philadelphia: W. B. Saunders.
Whitney, J. C. (1967). The pathology of the canine genital tract in false pregnancy. Journal of Small Animal Practice, 8, 247-263.
Zarrow, M. X., Sawin, P. B., Ross, S., Denenberg, V. H., Crary, D. D., Wilson, E. D., & Farooq, A. (1961). Maternal behavior in the rabbit: Evidence for an endocrine basis of maternal-nest building and additional data on maternal-nest building in the Dutch belted race. Journal of Reproduction and Fertility, 2, 152-162.
Zarrow, M. X., Yochim, J. M., & McCarthy, J. L. (1964).
The progestogens. In Experimental endocrinology: A sourcebook of basic
techniques (pp. 65-108). New York: Academic Press.