Physiology & Behavior, Vol. 50,
pp. 451-456. Pergamon Press plc. 1991.
Maintenance and Decline of the Suppression
of Infanticide in Mother Rats
L. C. PETERS,1 T. C. SIST AND M. B. KRISTAL2
Department of Psychology, State University of New York
at Buffalo, Buffalo, NY 14260
Received 16 November 1990
PETERS, L. C., T. C. SIST AND M. B. KRISTAL. Maintenance and decline
of the suppression of infanticide in mother rats. PHYSIOL BEHAV 50(2)
451-456, 1991. -- Virgin female rats kill foster neonates, whereas newly
parturient mothers do not. We demonstrated previously that this tendency
to kill is suppressed shortly prepartum, presumably by physiological factors.
In this study, we show that a) suppression of infanticide is maintained
through the first two weeks of lactation; b) the mothers that do not kill
foster neonates are not necessarily the same mothers that respond maternally
toward older foster pups, and those that kill neonates are not necessarily
the same ones that are nonmaternal to older pups, the two behaviors being
somewhat independent; and c) some virgins can be induced to be noninfanticides
by prolonged exposure to young, but only under special testing conditions
not required by actual mothers, which are nonkillers of foster young. This
suggests that the maintenance of the suppression of infanticide in mothers
owes something so the special circumstances of lactation other than continued
exposure to young.
Maternal behavior
Infanticide
Filicide Rats
Delivery
Lactation
Cannibalism
THE behavior of maternal rats includes pup-directed caretaking behaviors
such as retrieving of young to the nest, cleaning the young, and a specific
hovering posture that warms the pups and allows them to nurse (25,26).
In addition to caring for their young actively, it is essential that mother
rats not harm them. Although virgin rats generally kill freshly delivered
young that are still covered with fetal fluids and membranes, mothers given
such foster young very seldom kill (16, 21, 27). This indicates that infanticide
is somehow suppressed by the time rats become mothers. Like caretaking
behavior, the suppression of infanticidal responses first appears within
a day or so before delivery in primigravid rats. However, some rats that
show a suppression of their infanticidal tendencies when tested before
delivery do not actively care for foster pups. Other near-term pregnant
rats do actively care for older, cleaned foster pups, but will kill any
freshly delivered young they are given (21).
It is not clear how long the suppression of infanticidal tendencies
lasts. In laboratory tests of Long-Evans rats, it generally seems that
only hours-old neonates, uncleaned of fetal fluids and membranes, are killed
by virgin females (21). Is the suppression of infanticidal tendencies maintained
only for a day or so after birth? Alternatively, does it ever decline?
Various patterns have been seen in other rodent species. A male gerbil
kills any young it is given until some time during its mate's first pregnancy,
and thenceforth never kills again, whereas female gerbils refrain from
killing foster young only when they have young of their own (5). In at
least some environments, female hamsters kill a proportion of their own
litters, even after investing several days of care in them (3). In the
present experiments, we a) studied the duration of the suppression of infanticide
in mother rats; b) investigated the mechanisms by which it is suppressed;
and c) explored the degree to which that suppression is independent of
caretaking behavior.
GENERAL METHOD
The details of the method described here briefly can be found in Peters
and Kristal (21).
Subjects
Subjects were 170 adult female Long-Evans hooded rats kept on a 14-10
h light-dark schedule, with lights on at 0600 (EST). No rats other than
those indicated in Experiment 2 were tested more than once. Prior to testing,
all subjects were screened for estrous cyclicity and for spontaneous attraction
to placenta.
Apparatus
Tests were conducted in clear, wire-topped plastic cages, 21 x
42 x 20 cm. Minimal bedding was used so that
stimulus pups could not be buried or dried out by it.
Stimulus Pups
Long-Evans pups were used for tests of infanticide and maternal responsiveness.
Those used in infanticide tests were obtained 3 h before use by cesarean
section (hereafter referred to as "cesarean pups" or neonates) of donor
rats on Day 21 of gestation. Fetal fluids and membranes were allowed to
remain on the pups and were reconstituted with saline before testing. Placentas
were removed, however, so that placentophages and nonplacentophages would
not react differently to the stimulus pups (12,13). In an effort to exclude
moribund cesarean pups, we monitored the amplitude of their vocalizations
on an oscilloscope attached to a Holgate Mk IV ultrasonic receiver. Only
those pups were used that had responded with a minimum deflection in response
to 10 s of prodding with a cotton swab.
Procedure
Before rats were tested for infanticide, there was a 5-h period of habituation
to the test cage beginning at 1200 h. At 1700 h, the 60-mm infanticide
test was begun by placing a single cesarean pup (to keep the number of
pups exposed to harm to a minimum) into the end of the test chamber opposite
the corner where the subject slept or lay. Sixty mm after the stimulus
pups were presented, the experimenter returned and determined whether each
subject was a pup killer or a nonkiller.
EXPERIMENT 1: THE SUPPRESSION OF INFANTICIDE DURING LACTATION
AND AFTER WEANING
The first experiment was designed to determine how long after delivery
mother rats continue not to kill neonates. We tested a group of rats in
the second week of lactation [when suppression toward strange conspecifics
is most intense (6)], a group that was near the end of lactation, and a
group whose young had been weaned two weeks earlier. The behavior of these
groups was compared to that of large groups of virgins and newly delivered
mothers that also figured in a previous study (21), although, in many cases,
those rats were tested at the same time and in the same place as the rats
first discussed here.
METHOD
Subjects
Group L9 consisted of 17 rats tested on Days 8, 9, 10 or 11 of lactation
(Day 8, n = l; Day 9, n = l0; Day 10, n = 4; Day 11, n = 2). Group L20
was 10 rats tested on Days 19, 20 or 21 of lactation (Day 19, n = 4; Day
20, n = 4; Day 21, n = 2). Group NLP was 10 nonlactating primiparae tested
14 days after they had been separated from their 25- or 26-day-old litters
and had resumed normal estrous cyclicity as determined by vaginal smears.
Group Virgin was 52 virgins, and Group LI was 20 mothers tested on the
first full day of lactation. All lactating subjects were primiparous.
Procedure
Subjects were tested for infanticide in the manner described above.
During the 5-h period of habituation to their test cages, the lactating
rats had no contact with pups. Ten minutes after the end of the infanticide
test, most of the subjects in Groups L9, L20 and NLP were tested for 60
min for caretaking responses (retrieving, licking, crouching) toward 3
foster pups, 3-10 days old, placed in the end of the test cage opposite
the sleeping corner. In this way, it was possible to see if a rat was maternally
responsive even if it had killed a cesarean pup.
RESULTS AND DISCUSSION
Proportions of groups were compared using the standard error of the
difference between two proportions (4) because of the combination of large
n's and low expected cell frequencies.
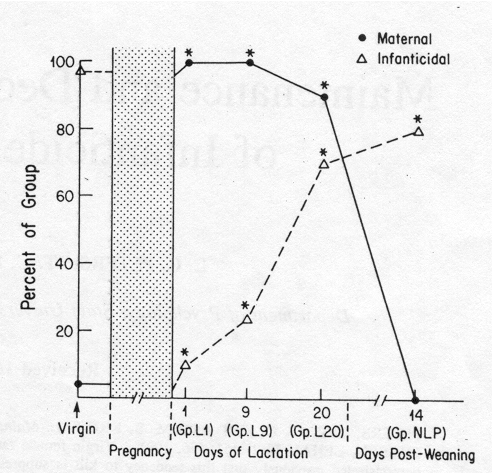
FIG. 1. Incidence of infanticide (toward newly cesarean-delivered pups)
and maternal behavior (toward normally delivered 3-day-old pups) among
groups of lactating primiparous rats. Values for pregnant rats can be obtained
from Peters and Kristal (21). (*Significantly different from virgins, p
< 0.05.)
Compared to virgins, significantly smaller proportions of primiparous mothers
killed cesarean pups not only on the first day after delivery (Group Virgin:
50 of 52 killed; Group L1: 2 of 20 killed; s
p1-p2
= 0.12; z = 7.3, p < 0.001), but also in the second week of lactation
(Group L9: 4 of 17 killed; s
p1-p2 =
0.12; z = 6.3, p < 0.001), near the end of lactation (Group L20: 7 of
10 killed, s p1-p2 = 0 .09;
z = 2.78, p = 0.003), and even two weeks after weaning (Group NLP: 8 of
10 killed; s p1-p2 = 0.08; z = 1.90,
p = 0.029). In Fig. 1, it can also be seen that, although the suppression
of infanticidal tendencies was maintained in some rats through lactation
and beyond, larger proportions killed as the interval after delivery increased.
Maternal responsiveness to older pups followed a similar pat tern, as
shown in Fig. 1. All 12 Group L9 rats given older pups responded maternally
to them. Nine of the 10 Group L20 rats given older (3-10 days old) pups
responded maternally, even though their own pups were much older (19-21
days old). None of the 8 Group NLP rats given older pups responded maternally
to them.
It appears that the suppression of infanticide is, in fact, maintained,
as caretaking is maintained, well beyond the first day or so when the hormonal
conditions are present that cause rapid induction of caretaking (30). Infanticide
is first suppressed prepartum, presumably due to hormonal or other physiological
changes of pregnancy, since the females in which this was seen had neither
undergone delivery nor had received previous exposure to young (21). That
the suppression of infanticide is maintained well into the lactation period
indicates one of two things:
(a) If suppression of infanticide is maintained by the same mechanism
that causes the onset of the suppression of infanticide prepartum, then
the mechanism must be different from the short-lived physiological basis
for the onset of caretaking behavior. The suppression would be based on
a physiological state that is present both shortly prepartum and through
much of lactation, e.g., the secretion of oxytocin (14,15) or prolactin;
or (b) if the suppression of infanticide is maintained through lactation
by a mechanism different from that responsible for its onset prepartum,
then it might be maintained either by physiological conditions specific
to the lactation period or else by nonphysiologic variables, such as experience
with young. Experience with young seems to be the principal basis for the
maintenance of caretaking behavior through lactation (26). In the following
experiments, we examined the effect of previous contact with young on rats'
infanticidal tendencies in an attempt to understand the factors that maintain
the suppression of infanticide through at least the first two weeks of
lactation and why infanticidal tendencies reappear as the time for weaning
approaches.
EXPERIMENT 2: THE EFFECT OF REPEATED EXPOSURE OF
VIRGINS TO NEONATES
Several reports have suggested that rodents that kill young may stop doing
so if repeatedly exposed to young (10, 18-20). Hersher, Richmond and Moore
(9) showed that when lactating sheep and goats were exposed for a time
to foster young that were protected from their attack, the adults would
then accept the young and not butt at them. The present experiment was
de signed to determine whether repeated or prolonged exposure to neonates
can suppress infanticide in virgins. If so, this would suggest that lactating
mothers continue not to kill infants because of habituation to them during
the first hours after delivery, when physiological conditions prevent mothers
from killing their own young.
METHOD
Subjects
Twenty-six pup-naive virgin rats were tested. Group NO- TOUCH rats (n
= 10) were exposed to cesarean pups presented in protective wire cubes.
Group TOUCH rats (n = 11) were ex posed to unprotected cesarean pups. A
Control Group (n = 5) consisted of subjects exposed to empty cubes.
Apparatus
Group TOUCH rats were individually housed and tested in 10-gallon glass
tanks. Group NO-TOUCH and Control rats were housed during the 7-day exposure
period in galvanized wire animal cages, 24.5 x
18 x 18 cm. All rats had lab chow and
water available ad lib. Small cages were used to decrease the likelihood
that the subjects could avoid the protected pups, since Terkel and Rosenblatt
(29) demonstrated that virgins become maternal more rapidly when they are
housed with pups in small cages.
The cesarean pups were protected from adults by two layers of screen.
The inner layer consisted of a pouch made from brass mesh (2 mm square)
that was large enough to allow the enclosed pup some movement. The outer
layer was a 7-cm cube made from strong galvanized mesh (1 cm square) that
opened on one side so the inner sleeve could be removed. Herrenkohl and
Lisk (8) have shown that virgins can be sensitized, although more slowly,
with only the exteroceptive cues from pups behind a wire screen.
Procedure
At 1200 h on the first day of testing, the subjects exposed to unprotected
pups (Group TOUCH) were moved into the glass tanks. Minimal bedding was
provided, making it difficult to bury pups or for the amniotic fluid on
the pups to be dried off. One 60-min infanticide test was conducted at
1700 h on each of six consecutive days.
The day before exposure to neonates began, NO-TOUCH and Control subjects
were moved to their small test cages and presented with empty wire cubes.
The following day, at about 1700 h, the cubes were removed and those of
the NO-TOUCH Group were replaced with cubes containing cesarean pups prepared
in the usual way, except that the membranes covering them had not been
moistened (1 pup/cube/subject). The Control subjects, which were not housed
in the same room as the experimental group, had their cubes returned empty
daily. This procedure was repeated with each member of Group NO-TOUCH receiving
a fresh cesarean pup daily until it had been exposed to pups for nearly
7 full days. At 1200 h on the day after the seventh presentation, both
the experimental and control groups were moved from their metal cages to
plastic test cages, and at 1700 h were given the infanticide test. At the
end of the test, subjects were given a 60-mm test for maternal responsiveness
to 3- to 10-day-old pups. Afterward, vaginal smears were obtained to determine
the subjects' ovarian cycle stages.
RESULTS AND DISCUSSION
All 11 subjects in Group TOUCH killed pups on every one of the six test
days. There seems to have been a trend for virgins to approach and kill
cesarean pups with shorter latencies as they acquired more experience with
them, although the particular data that would demonstrate this conclusively
were not systematically recorded. Apparently, if repeated exposure to neonates
has any suppressing effect on infanticide, it is overridden by some effect
of killing experience that promotes further killing. Therefore, the data
obtained from the NO-TOUCH Group were important in order to ascertain if
the development of suppression of infanticidal tendencies was possible
while avoiding any effects of killing experience.
Even after prolonged exposure to protected pups, Group NO- TOUCH did
not differ from the Control Group; all 15 rats killed cesarean pups. Moreover,
none of the rats was maternal to older pups.
The amount or type of pup contact in this experiment may not have been
sufficient to produce a suppression of infanticidal tendencies in experimental
subjects. In particular, the exteroceptive stimuli to which Group NO-TOUCH
was exposed may not have been adequate. In the subsequent experiment, attempts
were made to intensify the contact that virgins had with young. In this
way, we hoped to make a fair test of whether experience with newborn pups
can be sufficient to cause the infanticidal tendencies of virgins to be
suppressed to an extent comparable to that of mothers in the second week
of lactation.
EXPERIMENT 3: THE EFFECT OF PROLONGED - EXPOSURE
TO OLDER PUPS
Rosenblatt (23) and many others have shown that virgin female rats exposed
for several days to foster young (replaced daily with fed young) eventually
begin to care for these young in a way that is difficult to distinguish
from that of lactating mothers. This process of behavioral induction by
exposure is generally referred to as "sensitization." In this experiment,
we used sensitization as an especially intense form of exposure to young
(since it involves actual caretaking of young) to evaluate the role of
experiential factors in the suppression of infanticide during lactation.
If virgin females, i.e., females that lack the special physiological characteristics
of lactating mothers, can be induced by long-term exposure to young (and
by experience with not killing young) to be noninfanticides, then it would
be unnecessary to invoke hormonal mechanisms to explain the maintenance
of the suppression of infanticide for several days after delivery.
METHOD
Subjects
Thirty-five sensitized (maternal) virgin rats were used. Group NOPUP
consisted of 10 subjects tested for infanticide without older pups present
after a 5-h habituation period in which no pups were present. Group PUP
consisted of 10 subjects tested with 3- to 10-day old stimulus pups present
during both the 5-h habituation period and the infanticide test. The older
pups were allowed to remain with the subjects because the hormonal condition
of sensitized virgins has been shown to vary with the presence or absence
of pups (11). Group CUBE consisted of 10 subjects exposed to empty cubes.
The Control Group consisted of 5 subjects exposed to empty cubes. Group
CUBE and the Control Group were also tested with the older stimulus pups
still present during test-cage habituation and the infanticide test.
Procedure
Subjects had been induced to behave maternally (sensitized) by presenting
each with a stimulus litter of three or four pups 3 to 10 days old. After
each subject spent 24 h with a stimulus litter, the litter was returned
to its mother and replaced with a fresh one. Sensitization was judged to
have occurred when a subject retrieved, licked, and hovered over its pups
within 15 min after they were removed from the nest and scattered about
the cage. After the rat had met these criteria on 2 consecutive days, it
was considered eligible for inclusion in the experiment. Assignments to
groups were made such that subjects in the pairs NOPUP/PUP and CUBE/Control
each had taken about the same number of days to sensitize.
Wire cubes (each containing either a fresh cesarean pup or no pup) were
hooked tightly under the wire tops of the plastic cages as in Experiment
2. They were removed daily at around 1700 h. At about the same time, each
subject's group of three older stimulus pups was removed and replaced with
another that had been fed. At 1130 h on the fourth through seventh days
of exposure to cesarean pups, each subject's older pups were re moved from
the nest and scattered at the opposite end of the cage.
At 1200 h on the seventh day of exposure to cubes, all subjects were
given the infanticide test in fresh plastic cages. Three fresh 3- to 10-day-old
pups, without cubes, were also placed into each PUP, CUBE and Control test
cage. At the end of the infanticide test, noninfanticides were observed
for 5 mm to discover whether they were maternal to their (unprotected)
cesarean pups. The attractiveness of cesarean pups relative to older pups
for each noninfanticidal subject was then judged by scattering all four
pups at the end of the cage opposite the nest and observing the latency
to respond maternally to each pup. At the completion of testing, vaginal
smears were obtained from all subjects.
RESULTS AND DISCUSSION
Contact with older pups that was more than sufficient to induce maternal
behavior in virgins did not significantly reduce the proportion of subjects
killing neonates. Despite all the experience they received with older pups,
all 10 rats in Group NOPUP and 7 of 10 in Group PUP killed neonates (p
= 0.105, Fisher exact probability).
Infanticide was not inconsistent with maternal responsiveness, as at
least 6 of 7 of the Group PUP infanticides continued to treat the older
pups maternally during the infanticide test, whereas 2 of 3 noninfanticides
were maternal toward both kinds of pup (the remaining rat did not respond
maternally to either). No attempts were made to determine if Group NOPUP
rats were maternal during or after the test with cesarean pups, although
all of them were maternal on the morning before the test.
TABLE 1
PERCENTAGES OF INFANTICIDES AMONG GROUPS OF VIRGIN FEMALE RATS EXPOSED
TO PUPS
|
Experiment
|
Group
|
n
|
Condition
|
% Killers
|
| Experiment 2 |
NO-TOUCH |
10
|
Naïve Virgin |
100
|
|
TOUCH |
11
|
Naïve Virgin |
100
|
|
Control |
5
|
Naïve Virgin |
100
|
| Experiment 3 |
NOPUP |
10
|
Maternal Virgin |
100
|
|
PUP* |
10
|
Maternal Virgin |
70
|
|
CUBE* |
10
|
Maternal Virgin |
70
|
|
Control |
5
|
Maternal Virgin |
40
|
|
|
|
|
|
*Older pups remained
with subject during the test for infanticide of neonates.
Significantly different from Control, p<0.05. |
After 7 days of exposure to pups protected by cubes, 7 of 10
Group CUBE subjects killed cesarean pups. That infanticide was suppressed
in three subjects does not seem to be due to the 7 days of contact with
protected neonates, however, since only 2 of 5 Control rats killed (Table
1). The proportions of infanticides in the CUBE (70%) and Control (40%)
Groups did not differ statistically (p = 0.29,. Fisher exact probability).
All rats in both the CUBE. and Control Groups continued to respond maternally
toward older pups, but there was surprisingly little maternal responsiveness
toward cesarean pups. All 9 noninfanticidal sensitized virgins (3 from
Group PUP, 3 from Group CUBE, and 3 Controls) had been continuously housed
with older pups during their infanticide tests. By the end of those tests,
most of the cesarean pups had been retrieved and all had been at least
partly cleaned. Only in 2 of 9 cages, however, were neonates in a nest
with the three older pups. The differences in the treatment of older pups
and neonates were best seen in the retrieval test in which maternal responses
were noted during the 15 mm after the older pups and cesarean pups (by
then clean and dry) were scattered about the end of the cage opposite the
nest. Eight of the 9 noninfanticidal virgins retrieved all three older
pups to their nests, then licked and crouched over them, but only one retrieved
its cesarean pup to the nest (p = 0.0017, Fisher exact probability).
Two other noninfanticides retrieved cesarean pups to sites other than their
nests, licked them briefly, and then returned without them to crouch over
the older pups. None of the three that retrieved its cesarean pup did so
before all older pups. had been retrieved and licked. It is interesting
that, although neonates seemed to be inferior stimuli for maternal care
in these sensitized virgins, the virgins nevertheless did not kill them.
Of maternal virgins tested for infanticide while still in the presence
of older pups (Group PUP, Group CUBE and Controls), only 16 of 25 (64%)
killed cesarean pups. This proportion was clearly lower than that of virgins
in Experiment 1 that had not been given periods of prolonged exposure to
pups (50 of 52, or 96%) (s p1-p2
= 0.08; z= 3.77, p < 0.0002).
The proportion of infanticides among maternal virgins with pups present,
Groups PUP, CUBE and Control (64%), was significantly higher than that
found among Group L9 lactating mothers (24%) in Experiment 1 (s
p1-p2
= 0.16: z = 2.58; p = 0.005).
Although only 16 of 25 (64%) maternal virgins that had been allowed
to remain with their pups killed cesarean pups, all 10 Group NOPUP maternal
virgins killed cesarean pups after they had been separated from their pups
for 5 h (s p1-p2 = 0.16; z = 2.20;
p = 0.014).
There was no indication that the rats that killed tended to be in a
different stage of the estrous cycle than those that did not kill.
GENERAL DISCUSSION
We found in Experiment 1 that most mother rats still did not kill foster
cesarean pups in the second week of lactation. Experiment 2 demonstrated
that infanticide was not suppressed if killers were repeatedly exposed
to unprotected, freshly delivered pups. There also seems to have been no
inhibition of infanticide brought. about by prolonged exposure of virgins
to neonates housed in wire cubes. This was so when this was the only experimental
treatment (Experiment 2) and also when it was combined with the induction
of maternal behavior and continued exposure to older pups (Experiment 3).
In Experiment 3, we found that more intense exposure to older pups did
result in a significant reduction in the proportion of virgin infanticides
(from 100% to 64%), provided that older pups were present during both the
habituation and test periods.
It is still necessary to conclude that something besides pup contact
contributes to suppression of infanticide in mother rats. The noninfanticidal
virgins differed from noninfanticidal lactating mothers. All infanticide
tests given to mothers followed a 5-h period in which the mothers had no
contact with pups. Despite the separation, most mothers did not kill foster
neonates, whether they were in the first day of lactation (L1, when 10%
killed) or in the second week of lactation (L9, when 24% killed). The suppression
of infanticide in maternal virgins, however, in no case endured a 5-h separation
from the pups that produced it. Virgin and mother noninfanticides differ
in that only the latter are capable of withholding killing responses after
a 5-h separation from pups.
Some conclusions can be drawn about why mother rats continue to be noninfanticides
through much of their lactation period. First, contact with pups does appear
to play a role. The proportion of maternal virgins that did not kill stimulus
pups in the third experiment (9 of 25, or 36%) might have been even greater
if the subjects had been sensitized with pups that were younger, and hence
more like the neonates used in the infanticide test. It is unclear whether
it is necessary for virgins to be maternally responsive for pup contact
to turn them into noninfanticides. In any case, it seems likely that the
pup contact that mothers receive while being maternal is involved in maintaining
the suppression of infanticide -- infanticide that is first suppressed
prepartum by a physiological change, presumably hormonal, associated with
pregnancy (21).
Second, because mothers are affected little or not at all by a 5-h separation,
whereas infanticide-suppressed virgins are, other factors may be involved
besides pup contact. Mother rats differ from virgins in both a) the physiological
modifications they have undergone as a consequence of lactation, and b)
their history of having experienced parturition, including exposure to
neonates during a period when infanticide was suppressed. Either or both
of these differences could be responsible for the behavioral differences
of mothers and sensitized virgins.
If physiological aspects of lactation are involved in suppressing infanticide,
they may be the same physiological factors that appear to be responsible
for initiating the suppression of infanticide prepartum. At least two hormones,
prolactin and oxytocin, that are necessary for lactation and that are secreted
in response to suckling and other pup stimuli, are also secreted in the
hours immediately before parturition (7,17). It may be relevant that prolactin,
at least, is also present at greater concentrations in virgins that have
been sensitized than in those that have not been exposed to pups long enough
to become sensitized (11,28). However, the increase in prolactin levels
that is seen in sensitized virgins after reunions with their pups is only
a fraction of the increase seen in lactating mothers. This opens up the
possibility that sensitized virgins are not as resolutely noninfanticidal
as are mothers with pups of the same age because the latter have a more
highly developed neuroendocrine response to young (14,15).
It is also possible that mothers acquire a habit of suppressing infanticide
during the onset phase of the suppression. Unlike the sensitized virgins
in Experiment 3, mothers in the second week of lactation (L9) in Experiment
1 had the experience of not killing neonates before they underwent infanticide
tests. In the period shortly after their own parturitions, they may have
learned to respond to neonates in ways other than killing, and these habits
may have influenced their responses to the cesarean pups with which they
were subsequently tested. A similar long-term strengthening of maternal
caretaking by pup contact specifically during the first few hours after
delivery has been demonstrated by Bridges (1,2). Perhaps the nonkilling
by two of ten rats in Group NLP (the nonlactating primiparae of Experiment
1) was an expression of a long-term change in behavior not based in hormonal
conditions or recent exposure to young.
1Data were collected
by L.C.P. as part of a dissertation submitted in partial fulfillment of
the requirements for a Ph.D. degree at the State University of New York
at Buffalo. Now at the Department of Psychiatry. New York University Medical
Center.
2 Requests for reprints should be addressed
to Dr. M. B. Kristal, Department of Psychology, SUNY at Buffalo, Buffalo,
NY 14260.
REFERENCES
-
Bridges, R. S. Long-term effects of pregnancy and parturition upon maternal
responsiveness in the rat. Physiol. Behav. 14:245-249; 1975.
-
Bridges, R. S. Parturition: Its role in the long term retention of maternal
behavior in the rat. Physiol. Behav. 18:487-490; 1977.
-
Day, C. S. D.; Galef, B. G. Pup cannibalism: One aspect of maternal behavior
in golden hamsters. J. Comp. Physiol. Psychol. 91: 1179-1189; 1977.
-
Dixon, W. J.; Massey, F. J. Introduction to statistical analysis. New York:
McGraw-Hill; 1969.
-
Elwood, R. W. Changes in the responses of male and female gerbils (Meriones
unguiculatus) towards test pups during the pregnancy of the female.
Anim. Behav. 25:46-51; 1977.
-
Erskine, M. S.; Barfield, R. J.; Goldman, B. D. Intraspecific fighting
during late pregnancy and lactation in rats and effects of litter removal.
Behav. Biol. 23:206-218; 1978.
-
Grosvenor, C. E. Evidence that exteroceptive stimuli can release prolactin
from the pituitary gland of the lactating rat. Endocrinology 76:340-342;
1965.
-
Herrenkohl, L. R.; Lisk, R. D. The effects of sensitization and social
isolation on maternal behavior in the virgin rat. Physiol. Behav. 11:619-624;
1973.
-
Hersher, L.; Richmond, J. R.; Moore, A. U. Modifiability of the critical
period for the development of maternal behavior in sheep and goats. Behavior
20:311-320; 1963.
-
Jakubowski, M.; Terkel, J. Transition from pup killing to parental behavior
in male and virgin female albino rats. Physiol. Behav. 34: 683-686; 1985.
-
Koranyi, L.; Phelps, C. P.; Sawyer, C. H. Changes in serum prolactin and
corticosterone in induced maternal behavior in rats. Physiol. Behav. 18:287-292;
1977.
-
Kristal, M. B. Placentophagia: A biobehavioral enigma (or De gustibus
non disputandum est). Neurosci. Biobehav. Rev. 4:141-150; 1980.
-
Kristal, M. B.; Whitney, J. F.; Peters, L. C. Placenta on pups' skin accelerates
onset of maternal behavior in nonpregnant rats. Anim. Behav. 29:81-85;
1981.
-
McCarthy, M. M. Oxytocin inhibits infanticide in female house mice (Mus
domesticus). Horm. Behav. 24:365-375; 1990.
-
McCarthy, M. M.; Bare, J. E.; vom Saal, F. S. Infanticide and parental
behavior in wild female house mice: Effects of ovariectomy, adrenalectomy
and administration of oxytocin and prostaglandin F2a
.
Physiol. Behav. 36:17-23; 1986.
-
Mennella, J. A.; Moltz, H. Pheromonal emission by pregnant rats protects
against infanticide by nulliparous conspecifics. Physiol. Behav. 46:591-595;
1989.
-
Morishige, W. K.; Pepe, G. J.; Rothchild, I. Serum luteinizing hormone,
prolactin and progesterone levels during pregnancy in the rat. Endocrinology
92:1527-1530; 1973.
-
Noirot, E.; Richards, M. P. M. Maternal behaviour in virgin female golden
hamsters: Changes consequent upon initial contact with pups. Anim. Behav.
14:7-10; 1966.
-
Paul, L.; Herrenkohl, L.; Kupferschmidt, J.; Whitney, J. Conspecific young:
The ability of the same stimulus complex to induce both cannibalism and
maternal behavior. Paper presented at the Eastern Conference on Reproductive
Behavior, Nags Head, NC; June 1975.
-
Paul, L.; Kupferschmidt, J. Killing of conspecific and mouse young by male
rats. J. Comp. Physiol. Psychol. 88:755-763; 1975.
-
Peters, L. C.; Kristal, M. B. Suppression of infanticide in mother rats.
J. Comp. Psychol. 97:167-177; 1983.
-
Reisbick, S.; Rosenblatt, J. S.; Mayer, A. D. Decline of maternal behavior
in the virgin and lactating rat. J. Comp. Physiol. Psychol. 89:722-732;
1975.
-
Rosenblatt, J. S. Nonhormonal basis of maternal behavior in the
rat. Science 156:1512-1514; 1967.
-
Rosenblatt, J. S. Views on the onset and maintenance of maternal behavior
in the rat. In: Aronson, L. R.; Tobach, E.; Lehrman, D. S.; Rosenblatt,
J. S., eds. Development and evolution of behavior. San Francisco: W. H.
Freeman and Co.; 1970.
-
Rosenblatt, J. S.; Lehrman, D. S. Maternal behavior in the laboratory rat.
In: Rheingold, H. L., ed.. Maternal behavior in mammals. New York: Wiley;
1963.
-
Rosenblatt, J. S.; Siegel, H. I.; Mayer, A. D. Progress in the study of
maternal behavior in the rat: Hormonal, nonhormonal, sensory and developmental
aspects. Adv. Stud. Behav. 10:225-311; 1979.
-
Stern, J. M. Parturition influences initial pup preferences at later onset
of maternal behavior in primiparous rats. Physiol. Behav. 35: 25-31; 1985.
-
Stern, J. M.; Siegel, H. I. Prolactin release in lactating primiparous
and multiparous thelectomized and maternal virgin, rats exposed to pup
stimuli. Biol. Reprod. 19:177-182; 1978.
-
Terkel, J.; Rosenblatt, J. S. Aspects of nonhormonal maternal behavior
in the rat. Horm. Behav. 2:161-171; 1971.
-
Terkel, J.; Rosenblatt, J. S. Humoral factors underlying maternal behavior
at parturition: Cross transfusion between freely moving rats. J. Comp.
Physiol. Psychol. 80:365-371; 1972.