Suppression of Infanticide in Mother Rats
Lawrence C. Peters and Mark B. Kristal
State University of New York at Buffalo
In the present series of experiments, we (a) show how under controlled conditions virgins are much more likely than mothers to kill surgically delivered young, (b) show that the same is true when naturally delivered young are used as stimuli, (c) investigate the mechanisms by which infanticide becomes suppressed in mothers, and (d) study the relations of the onset of the suppression of infanticide to the onset of active caretaking behaviors.
General Method
Subjects
Subjects were 165 female Long-Evans hooded rats, at least 70 days old, which were kept on a 14:10 hr light/dark schedule, with lights on from 0600 to 2000 hours (EST). When not being tested, the rats were housed individually in galvanized wire mesh cages, 24.5 x 18 x 18 cm. Food (Charles River Rat/Mouse/ Hamster Formula 3000) and water were available ad lib. Some subjects (n = 25) were purchased from Charles River Breeding Laboratories; the rest were born in our laboratory to parents that had been purchased from Charles River. The purchased rats were used in several different experimental groups and did not appear to respond differently from the rats born in our laboratory. Beginning at about 60 days of age, subjects were screened for regular ovarian cyclicity (by daily inspection of vaginal smears) and were tested for spontaneous placentophagia according to the method described by Kristal and Graber (1976). Unless it is stated otherwise, no rat was tested for infanticide more than once. Except for rats in the "mother" groups, which were primiparous, all subjects were nulliparous.
Apparatus
Tests were conducted in clear, wire-topped plastic cages, 21 x 42 x 20 cm. Only small quantities of bedding were used so that stimulus pups could not be buried or dried out by it. Rats in the test room were able to hear and smell each other, but visual contact between them was blocked.
The amplitude of pup vocalizations was monitored with an oscilloscope which received input from a bat detector (Holgate Ultrasonic Receiver MK.V.). The bat detector was used primarily to amplify the signal and transmit it from the microphone to the oscilloscope. The microphone was suspended 7.5 cm above the stimulus pup.
Stimulus Pups
Pups used for infanticide tests and for tests of maternal responsiveness were obtained from Long- Evans donor females that had been bred to Long- Evans males. The day on which sperm was found in the vaginal smear of a donor was considered to be Day 1 of pregnancy. The pups used for infanticide tests either were obtained by cesarean section on Day 21 (Experiments 1 and 3) or were delivered normally on Day 22 or 23 (Experiment 2).
Procedure
On the day of testing, vaginal smears were obtained from all intact cycling subjects to determine the stage of the estrous cycle. Before the rats were tested for infanticide, there was a 5-hr habituation period beginning at 1200 hours when subjects were placed in clean test cages. At about 1630 hours, the end of the cage in which each rat was sleeping was noted. At 1700 hours, the 60-mm infanticide test began with the placing of a single newborn pup into the end of the test chamber opposite the sleeping corner. The stimulus pup was handled with a disposable plastic glove. The experimenter remained in the test room only for the first 5 mm. During that time a note was made of the latency for each subject to approach and sniff its stimulus pup. Sixty minutes after the stimulus pups were presented, the experimenter returned and determined whether each subject was a pup killer (if her pup was missing or showed signs of having been bitten) or a nonkiller. Very seldom did a pup killer not consume the entire pup during a 60-min test. To be designated as a nonkiller, a rat had to show some sign of having interacted with the pup as well as of not having harmed the pup. An interaction was said to have occurred if any of the following were observed or inferred: (a) sniffing the pup during the initial 5 min, (b) licking the pup, as indicated by the absence of membranes and fluid on the pup at 60 min, or (c) carrying the pup, as indicated by finding it in the end of the cage opposite the one where it was placed at the beginning of the test (cesarean pups did not locomote well enough to get that far on their own). Thus, rats were not called nonkillers if they were simply too afraid of pups (or conceivably had too little interest in them) to approach and interact with them during the test. Only five subjects were excluded because they were classified neither as pup killers nor as nonkillers.
Statistical Analysis
Since all of the infanticide and maternal behavior data
were nominal (e.g., either a rat was a pup killer or it was not), analyses
consisted of pairwise comparisons of group proportions. The statistical
test that was used for a particular comparison depended on group size and
expected cell frequency. To compare small groups (n < 20)
or medium-sized groups (20 < n < 40) with an
expected cell frequency less than five, we used Fisher's exact probability
test (Siegel, 1956). In other cases, z was calculated from the standard
error of the difference between two proportions (Dixon & Massey, 1969).
Experiment 1: Responses of Virgins and Mothers to Cesarean-Delivered Pups
Before we began investigating the phenomenon of the suppression of infanticide, it was first necessary to develop a method of testing rats for infanticidal tendencies that would provide us with unambiguous, repeatable results. Although Sachs (Note 2) did show that there were differences in the responses of virgin and mother rats to foster neonates, the differences were not large. Although many of his virgins killed pups, many did not. Some of the latter ate only placentas, although most did not disturb either pups or placentas. On the other hand, many of Sachs's mothers ate foster pups as well as placentas. Much of this variability in rats' responses to the cesarean-delivered pup-placenta unit can be explained by (a) variability in the viability of the pups and (b) the presence of placentas, connected by their umbilical cords to the pups. Kristal and Graber (1976) showed that most virgin rats presented with donor placenta not only do not eat it but actually seem to avoid it. Since virgins presented with pups and attached placentas might be expected to avoid both the pup and placenta if the placenta is aversive, we decided to use pups from which placentas had been removed.
The tendency of many mothers to kill and eat cesarean-delivered pups might be explained by the fact that the pups were moribund when the mothers were exposed to them. Sachs (Note 2) showed that female rats in all reproductive conditions are more likely to eat dead cesarean-delivered pups than live ones. Since most of Sachs's pups died before he was able to present them to subjects and since Paul et al. (Note 1) observed that many cesarean-delivered pups died spontaneously soon after they were given to mothers, it seems important to use only pups that are strong and healthy and not likely to die during an infanticide test unless they are killed by the rats being tested.
Method
Subjects. On the day after parturition (and after between 21 and 35 hr of experience with their own pups), 20 primiparous mothers were tested with cesarean-delivered pups that had not been cleaned of their fetal fluids and membranes but that had had their placentas removed (Group Mother; see Table 1). Their responses to these pups were compared with responses of 22 virgins that were tested with pups of the same type. Of these virgins, 15 were placentophages (Group PL-Phage), and 7 were nonplacentophages (Group NonPL-Phage). Screening for placentophagia had been conducted earlier according to the method described by Kristal and Graber (1976). Another group of 7 nonplacentophages (Group PL/NoPL) was tested with uncleaned pups whose placentas remained attached and then 2 days later was retested with pups whose placentas had been removed. In this way an assessment could be made of the effect of the presence or absence of placenta on infanticide in the same group of nonplacentophages.
Table 1
Groups in Experiment 1
|
|
|
|
|
| PL-Phage |
|
virgin placentophages | uncleaned, no placenta |
| NonPL-Phage |
|
virgin nonplacentophages | uncleaned, no placenta |
| Mated Virgin |
|
mated, not pregnant | uncleaned, no placenta |
| Combined Virgin |
|
first three groups combined | uncleaned, no placenta |
| PL/NoPL |
|
virgin nonplacentophages | uncleaned, placenta/no placenta |
| Cleaned Pup |
|
virgin placentophages | cleaned, no placenta |
| Mother |
|
mothers, first day of lactation | uncleaned, no placenta |
To determine how the presence or absence of fetal fluids and membranes affects the response of virgins to neonates, we presented 17 virgin placentophages (Group Cleaned Pup) with cesarean-delivered pups that had been cleaned with distilled water and that had had membranes, umbilical cord, and placenta removed.
As a control for the effects of mating and handling on response to pups, uncleaned cesarean-delivered pups without placentas were presented to a group of six placentophages and one nonplacentophage that had been mated and housed in cages adjacent to those of pregnant rats but had not themselves become pregnant. These females (Group Mated Virgin) were tested on what would have been Day 22 had they been pregnant.
Stimulus pups. Stimulus pups were taken from donor females by cesarean section at 1400 hours on Day 21 of pregnancy. These fetuses (henceforth called cesarean pups") were quickly removed from the uterus, along with their fetal membranes and umbilical cords. Working as rapidly as possible, we removed all material from the pups' breathing passages and then placed the pups on a paper towel in a covered earthen vessel warmed by a hot water bath kept between 35 and 37C. The first time that Group PL/NoPL was tested, placentas were left attached to the umbilical cords. The placentas, which dry out rapidly, were kept physically separate from the pups and associated fluids and membranes and were kept moist in saline-soaked gauze. In all other groups, placentas were detached from their umbilical cords shortly after the pups were delivered. At no time were the pups touched with bare hands. Both immediately after delivery and for several minutes after they were placed in the warming vessels, pups were lightly rubbed and prodded with dry cotton swabs, much as naturally delivered pups are vigorously licked and mouthed by their own mothers.
Despite the care that was taken, many of these pups died soon after they were delivered, just as did most of those that Sachs (Note 2) and Paul et al. (Note 1) delivered by cesarean section. If, however, a pup could survive for 3 hr after delivery (62% did), then it was almost certain to survive for at least several more hours. For this reason we performed cesarean deliveries 3 hr before the infanticide test.
Shortly before we used stimulus pups, we measured the amplitude of their vocalizations in response to 10 sec of prodding with a cotton swab. Those that could not produce at least a 1-mV deflection of the oscilloscope beam were not used. Whether or not pup vocalizations play a role in inhibiting infanticide in mothers, the intensity of tactually elicited vocalization did appear to be related to the intensity of other behavioral responses to handling. Pups that met the criterion for vocalization were capable of a great deal of movement and vocalization even if they were kept at room temperature for several hours.
After we measured vocalization amplitudes, and immediately before we began testing, we moistened each pup with three drops of .9% saline. Thus the fetal membranes and amnionic fluid that had dried on the pups were more or less reconstituted. During the moistening procedure, the tray containing the pups was placed in the 35-37°C water bath for 2 min.
Procedure. The testing procedure was that described in General Method.
Results
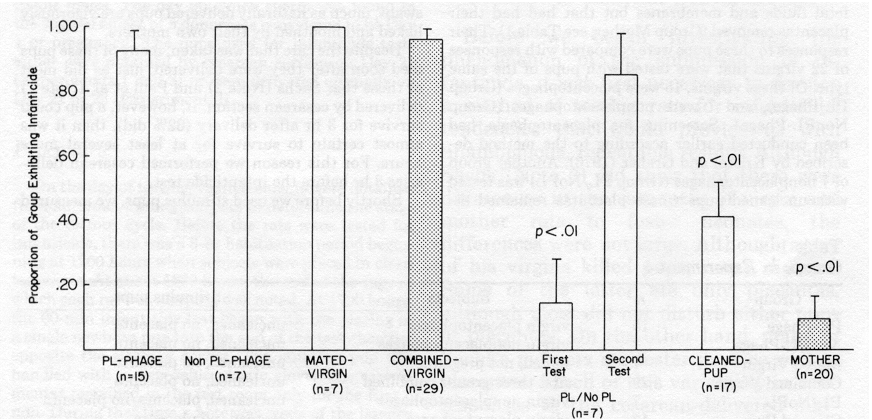
From Figure 1 it can be seen that virtually all virgins killed and ate uncleaned pups presented without placentas, whether the virgins had previously been shown to be placentophages or nonplacentophages. Since the subjects in Group Mated Virgin did not become pregnant and were presented with uncleaned pups without placentas, they were grouped together with unmated placentophages (Group PL Phage) and nonplacentophages (Group NonPL-Phage) given similar stimuli. The combination of these three groups was designated as "Group Combined Virgin," of which 28 of 29 females killed neonates.
Although nonplacentophages killed pups when placentas were absent, most did not kill pups when placentas were present. The first time Group PL/NoPL rats were tested, a significantly smaller proportion of nonplacentophages killed (1 of 7) than did in Group NonPL-Phage, which were given pups without placentas (7 of 7; p = .002).
Unlike virgin nonplacentophages given pups without placentas, most of Group PL/ NoPL did not approach and sniff their pups in the first 5 mm of the test, or even lick the fluids and membranes off their pups during the hour-long test with placenta pre sent. However, when these same rats were tested 2 days later with uncleaned pups that did not have placentas attached, all but one killed and ate the pup.
Virgins were less likely to kill pups that had been cleaned of fluids and membranes than they were to kill pups that had not been cleaned. A significantly smaller pro portion of Group Cleaned Pup (7 of 17) killed pups than of Group PL-Phage (14 of 15; z = 3.10, p = .013). Of the 10 virgins that did not kill cleaned pups, 7 were seen to approach and sniff their pups, and 4 were observed to lick them.
The most important finding was that the proportion of mothers that killed
uncleaned pups presented without placentas (2 of 20) was significantly
smaller than that of Group Combined Virgin given the same sort of stimulus
(28 of 29; z = 6.11, p < .001). Nearly all nonkiller
mothers licked the fluids and membranes from their pups and crouched over
them. This was so, even though the pups were not the mothers' own, having
been obtained from donor mothers.
Figure 1. Proportions (± SE) of groups of virgins and of mothers that exhibited infanticide. (Statistical comparisons are with Group Combined Virgin. PL = placenta.) No relation was found in any group between stage of the estrous cycle and out come of the infanticide test, either in this experiment or in others with cycling virgins (Peters & Kristal, Note 3).The rats that were observed to kill cesarean pups did so simply by picking them up and eating them, as they would a piece of food. In no case was a rat seen to attack a cesarean pup as adult rats sometimes at tack juveniles or mice, i.e., by springing upon them and biting them repeatedly be fore settling down to eat them (Paul & Kupferschmidt, 1975).
Discussion
Mother rats tested on the first full day of lactation are clearly less likely than are virgins to kill uncleaned cesarean pups presented without placentas. The difference would probably not be so clear if cleaned pups had been used, since virgins are apparently less likely to kill pups after the fluids and membranes have been removed. After the pups had been cleaned, virgins seemed not to regard them as food. On the other hand, rats that killed newborn pups seemed only to be eating them rather than attacking them.
Nonplacentophages consistently killed neonates, provided that they were presented without placentas. When the placenta was present, nonplacentophages did not seem even to approach the pups. This is not surprising, given the aversion that nonplacentophages seem to have for the placenta presented by itself (Kristal & Graber, 1976). Although known placentophages were not tested with uncleaned pups attached to placentas in this experiment, other work has shown that placentophages given uncleaned neonates and several placentas invariably eat both the pups and the placentas (Peters, 1980). Apparently placentas are aversive to virgin nonplacentophages and attractive to virgin placentophages. When the former become mothers, they become, for a time, avid placentophages; the presence of placentas does not keep them away from pups (Kristal, 1980).
This first experiment confirms Sachs's (Note 2) finding that when a
female rat becomes a mother, her tendency to kill uncleaned neonates is
suppressed. Not only does she not kill her own uncleaned infants when they
appear, but the inhibition of killing extends also to foster infants. This
experiment also makes it clear that the suppression of infanticidal tendencies
in mother rats is most easily seen when lively uncleaned neonates are used
and their placentas are absent. With this established, it was then necessary
to find out whether inhibition of the killing of foster pups is limited
to cases when cesarean-delivered Day 21 pups are used or whether virgins
and mothers also differ in their responses to full-term pups.
Experiment 2: Responses of Virgins and Mothers to Full-Term Pups
Compared with Day 21 cesarean pups, full-term naturally delivered pups, with which mothers interact at parturition, are larger and more mature. It is possible that natural delivery and an additional day of maturation in utero confer some important sensory quality on full-term pups that cesarean pups lack. To test this possibility, we compared the responses of virgins and mothers to naturally delivered pups.
Method
Subjects. Ten pup-naive virgins and nine primiparous mother rats in the first week of lactation were used as subjects.
Stimulus pups. Donors were placed in high-walled glass tanks (10-gal [.04-m3] aquaria) shortly before they were due to give birth. Immediately after delivery, pups were removed from the tanks with forceps before their mother had much opportunity to lick them and strip them of their membranes. Like cesarean pups in Experiment 1, naturally delivered pups were rubbed with cotton swabs after delivery, the amplitude of their vocalizations was measured, and immediately before testing, the pups were moistened with distilled water and warmed. Also, as with cesarean pups, these pups were presented with attached fetal membranes but without placentas.
Procedure. Without modifying the light cycle, it is not possible to predict more than an hour or two in advance at what time of day a rat will deliver, or even on which of the two usual days she will deliver (Mayer & Rosenblatt, 1980). For this reason we were unable to time events as exactly as when we used cesarean pups. However, the beginning of the test was confined to the period between 1500 and 1800 hours, habituation to the test cages lasted between 3 and 6 hr, and pups were used between 1 and 4 hr after their delivery. (Since naturally delivered full-term pups are generally quite vigorous at birth, it was not necessary to wait 3 hr before using them.) The reason for the range in pup ages (1-4 hr) is that puerperae become very emotional when their pups are removed (i.e., they vocalize and attempt to bite the experimenter), and they very often delay delivery of the next pup. The test was conducted in the same way as was described above for cesarean-pup tests.
Results and Discussion
Eight of the ten virgins killed, whereas only two of the nine lactating mothers killed (p = .019).
There was more variability in the tests with full-term pups than in the tests with cesarean pups: Test time and duration of the habituation period varied with the availability of stimulus pups. Since it is difficult to prevent donors from licking some of the fluid and membranes from pups as they are being delivered, the amount and kind of substances on the pups also varied from pup to pup. When cesarean-delivered pups are used, these problems can be avoided. Hence, subsequent experiments were done with cesarean pups.
To choose between these three explanations of the apparent suppression of infanticide in mothers, and also to examine the temporal relation between the offset of infanticide and the prepartum onset of maternal responsiveness, we used both neonates and older pups to test rats that had gone through pregnancy but not parturition. Testing them with newborn pups made it possible to tell whether changes connected with pregnancy are sufficient for infanticide to be suppressed. Testing the same individuals with somewhat older pups (which even "pup killers" would not kill) made it possible to discern whether maternal caretaking always occurs together with nonkilling or whether they develop independently of each other.
Method
Subjects. Two kinds of experimental treatment were used. One was to test intact pregnant rats shortly before delivery, and the other was to test rats after their pregnancies had been surgically terminated. Group Day 22/I-MR (n = 25) rats were tested on Day 22 of pregnancy, first for infanticide (I) with cesarean pups and then for maternal responsiveness (MR) with older pups (with which maternal responsiveness could be judged even if the neonates had been killed). They were compared with the 29 rats of Group Combined Virgin from Experiment 1, which had been tested with cesarean pups only. Group Day 22/MR-I (n = 13) consisted of pregnant rats tested in the opposite order: first with older pups and then with cesarean pups. They were compared with Group Virgin/MR-I (n = 15). The 12 rats in Group Day 23-H/I-MR were hysterectomized while still pregnant on Day 22, then tested on the next day in the I-MR order. Their control group was Group Virgin-H/I-MR (n = 5), consisting of virgins also hysterectomized and then tested on the next day. Unlike other rats in these experiments, the two hysterectomized groups were not pretested for their responses to placenta.
Surgery. Hysterectomy was performed through a single midventral incision under methoxyflurane anesthesia (Metofane, Pitman-Moore) on the morning of Day 22. The uterus was removed by cutting each horn immediately below the fallopian tube and by cutting the body of the uterus immediately above the cervix. Rats were ambulatory shortly after this operation and appeared to behave normally by the next day.
Procedure. Subjects that were to become pregnant were bred and then housed individually in wire mesh cages. The day before they were to be tested, they were moved to plastic cages with wood-shaving bedding. Those that delivered before they could be tested (11 of 49) were dropped from the study. No rats in Group Day 23-H/I-MR delivered before they could be hysterectomized.
Although the 60-mm infanticide test was always begun at 1700 hours, the 60-min test for maternal responsiveness was begun either after or before the infanticide test (depending on whether the subjects were in Group I-MR or Group MR-I). There was a 10-min interval between tests, during which time pups were absent. In the test for maternal responsiveness, three pups 3-10 days old were placed in the corner of the test cage opposite the subject's sleeping corner. The subject's responses to them were observed for 15 min and then checked at 15-min intervals until the end of the 60-min test. Subjects were considered to be maternally responsive to the older pups if they retrieved at least two, licked them, and then remained huddled over them.
Since subjects sometimes seemed to retrieve and lick neonates only because of the attached fluids and membranes (and ignored neonates once those materials were licked off), a rat was considered to have responded maternally to a cesarean pup only if the pup was retrieved, was licked, and continued to receive maternal attention at the end of a 60-min test. Continued maternal attention was indicated by the female's crouching over the pup, by warmth detected when the pup was picked up by the experimenter, or by continued (if only occasional) licking of the pup long after the fetal membranes and fluid had been cleaned from its skin.
After the tests for infanticide and maternal responsiveness were completed, each pregnant subject was returned to her plastic maternity cage and moved to another room. She was not disturbed again until after she had given birth. Frequent checks were made in order to determine the time at which the first pup was delivered. Rats that were not pregnant became Group Mated Virgin of Experiment 1.
The hysterectomized subjects were tested at the usual time on the day after surgery. Each test with cesarean pups was followed by a 60-mm test for maternal responsiveness to pups 3-10 days old.
Results and Discussion
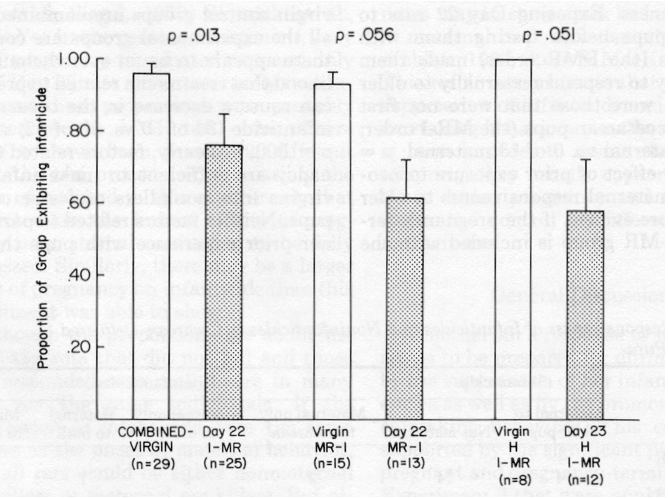
The proportion of each group that killed cesarean infants is shown in
Figure 2. Al though .97 (28 of 29) of Group Combined Virgin killed, a smaller
proportion, .76 (19 of 25), of Group Day 22/I-MR killed (z =
2.24, p = .013). Similarly, whereas .93 (14 of 15) of
the control rats of Group Virgin/MR-I killed, .62 (8 of 13) of Group Day
22/MR-I killed (p = .056). In the hysterectomized groups, 1.00 (8
of 8) of Virgin-H/I-MR killed, and .58 (7 of 12) of Day 23-H/I-MR killed
(p = .051).
The order in which rats were tested with cesarean pups and older pups did not affect their infanticide response to cesarean pups. All or nearly all virgins killed pups regard less of test order and regardless of whether they had been hysterectomized. Between .24 and .42 of each pregnant or pregnancy- terminated group were nonkillers.Figure 2. Proportions (± SE) of late pregnant, pregnancy-terminated, and virgin-control groups that exhibited infanticide. (I = infanticide; MR = maternal responsiveness; H = hysterectomized.)
The results of tests for maternal responsiveness are shown in Table 2 (results are reported only for the 23 rats in Group Day 22/I-MR that were tested with older pups). Only .31 of pregnant and pregnancy-terminated rats (15 of 48) met the criteria for maternal responsiveness toward older pups, cesarean pups, or both. Still, there were pup killers that were maternally responsive as well as those that were not; there were also nonkillers that were maternally responsive and those that were not. It is surprising that although test order did not affect the likelihood of infanticide, it did seem to affect the likelihood of maternal responsiveness. Exposing Day 22 rats to cesarean pups before testing them with older pups (the I-MR order) made them more likely to respond maternally to older pups than were those that were not first exposed to cesarean pups (the MR-I order; 6 of 23 maternal vs. 0 of 13 maternal; p = .052). The effect of prior exposure to neonates on maternal responsiveness to older pups is more evident if the pregnancy-terminated I-MR group is included with the Day 22 I-MR group (12 of 35 vs. 0 of 13; z = 2.44, p = .007). Of the 23 virgin controls that were tested with older pups as well as with neonates, only 1 (in Group Virgin-H/I-MR) was maternally responsive.
Table 2
Maternal Responsiveness of Infanticides and Noninfanticides to Cesarean-Delivered
and Older Pups
|
|
|
|||||||
| Group | Maternal to older pups | Not maternal | Maternal only to cesarean | Maternal only to older | Maternal to both | Maternal to neither | ||
| Day 22,
I/MR
(n = 23) |
4 | 14 | 2 | 1 | 1 | 1 | ||
| Day 22,
MR/I
(n = 13) |
0 | 8 | 1 | 0 | 0 | 4 | ||
| Day 23,
H-I/MR
(n = 12) |
3 | 4 | 0 | 1 | 2 | 2 | ||
Careful records were kept of the delivery times of rats tested on Day 22; there was no apparent relation between the length of the test-to-delivery interval and the proportion of subjects that had been infanticidal or maternally responsive.
Since prior exposure to older pups did not seem to change the responses of virgins or pregnant females to cesarean pups, the Day 22 I-MR and MR-I groups can be regarded as replications of the same treatment, at least in regard to infanticide. If the Day 22 groups are combined and their virgin control groups are combined, the difference between the infanticidal tendencies of prepartum Day 22 rats and virgins be comes more clear (27 of 38 vs. 42 of 44; z = 3.02, p = .001).
It is possible that if a different procedure had been followed, a smaller proportion of the pregnancy-terminated group would have been infanticidal. However, it is clear that after hysterectomy during pregnancy, as few as .58 of rats kill. Hence, if all the virgin control groups are combined and if all the experimental groups are combined, there appears to be an overwhelming likelihood that treatments related to pregnancy can cause a decrease in the occurrence of infanticide (34 of 50 vs. 50 of 52; z = 3.72, p = .0002). Clearly, factors related to pregnancy are sufficient to make infanticidal virgins into nonkillers of foster cesarean pups. Neither factors related to parturition nor prior experience with pups that they themselves have delivered is necessary to prevent mother rats from killing foster neonates. This finding strongly suggests that mother rats are nonkillers at delivery due to changes that occur during pregnancy. The question that remains is, Why was pregnancy sufficient to make nonkillers out of a third of the rats that were tested but not the other two thirds? If pregnancy is normally sufficient to suppress infanticide in only a third of mothers, then parturition or other factors may also be necessary for the suppression to occur in the rest of them.
It seems likely that factors related to pregnancy normally have more effect on maternal behavior than was seen in this experiment. There is a great deal of evidence indicating that virtually all rats be come maternally responsive when their pregnancies are experimentally terminated in the appropriate way (Rosenblatt, Siegel, & Mayer, 1979). In a study in which pregnancy was surgically terminated shortly before term, .88 of the rats responded maternally within 5 mm after foster pups were presented (Moltz, Robbins, & Parks, 1966). Other investigators who have looked for prepartum maternal behaviors have found them in most of the rats they tested (Rosenblatt & Siegel, 1975; Slotnick et al., 1973). Yet in the present experiment, only .50 of pregnancy-terminated rats and only .25 of Day 22 rats were maternally responsive, and those proportions were probably elevated by the I-MR test order. There seems to have been something different about the test procedures, or possibly the strain of rat used in this experiment, that did not allow the known effects of pregnancy on maternal caretaking to be fully expressed. Similarly, there may be a larger effect of pregnancy on infanticide than this experiment was able to show.
Although the proportions are about the same, the rats that did not kill and those that responded maternally were in many cases not the same individuals. If the suppression of infanticide were the same process as the onset of maternal behavior, then all rats would be either nonmaternal pup killers or maternal nonkillers. But almost a third of those tested (14 of 48) were either maternal pup killers or nonmaternal nonkillers (Table 3). These 14 rats were nearly two thirds of those in which pregnancy affected behavior (i.e., that were no longer nonmaternal pup killers like virgins). Differences in rats' sensitivities to pups of different ages are probably not responsible for this apparent independence of the suppression of infanticide from the onset of maternal responsiveness. Rats that were noninfanticidal were about as likely to respond maternally to older pups as to cesarean pups; a number of rats that killed cesarean pups also responded maternally to older pups despite indications that neonates may in some cases be better elicitors of maternal responsiveness (Mayer & Rosenblatt, 1980; Stern & MacKinnon, 1978). This result demonstrates that although the suppression of infanticide and the onset of caretaking behaviors must both be present at term in order to assure the survival of offspring, they usually appear at different times during pregnancy and are probably under the control of different biological mechanisms.
Table 3
Outcome of Tests for Infanticide and Maternal Responsiveness Among
Day 22 Pregnant and Pregnancy-Terminated Groups Combined
|
|
||
| Caretaking |
|
Noninfanticidal |
| Maternally responsive |
|
|
| Not maternally responsive |
|
|
|
Note. Forty-eight rats were tested for maternal responsiveness a Rats that behaved like mothers. b Rats that behaved like virgins. |
||
General Discussion
A mother rat's response to her own pups seems to be prepared for during prepartum by the suppression of her infanticidal tendencies as well as by the promotion of active caretaking behaviors. This conclusion is supported by the significant proportions of pregnant and pregnancy-terminated rats in Experiment 3 that were nonkillers in their first-ever exposure to neonates. Because of this we can rule out the possibility that the mothers in Experiments 1 and 2 did not kill foster infants only because of prior experience with not killing their own infants. Instead, the killing of either a rat's own or foster pups is prevented by changes during pregnancy that are at least partly independent of those that control the onset of maternal responsiveness.
The positive effect of prior exposure to uncleaned neonates on maternal responsiveness toward older pups may have some relevance to the ways in which maternal responsiveness normally becomes established at parturition. It may be that the fluids and membranes clinging to the neonates were responsible for the increased response. Nearly all rats, pup killers or not, consume those substances, and it was shown in Experiment 1 that their presence gets many rats orally involved with pups that otherwise they would not much attend to. Kristal, Whitney, and Peters (1981) found that attractive materials placed on the skin of older pups (3-8 days old) reduced the number of days of pup exposure required for virgins to become maternally responsive. The best such material, when the subjects were placentophages, was a combination of foster placenta and amniotic fluid.
In addition to the effect on maternal responsiveness of substances clinging
to their skin, infants may possess other qualities that make them more
attractive than older pups, at least in some situations. Untreated virgins
developed maternal responsiveness more quickly when they were given 1-2-day-old
pups than when the pups were 3 days old or older (Stern & MacKinnon,
1978). Mayer and Rosenblatt (1980) found that most hormonally treated virgins
responded maternally almost immediately to neonates taken from their mothers
within 2 hr after birth, although most hormonally treated virgins took
considerably longer to respond to 3-8-day-old pups. There may have been
some residual membrane and fluid on the skins of those neonates. How ever,
whether it is the fluids and membranes or whether it is something else
that is responsible for the attractiveness of neonates, that attractiveness
seems to have generalized to the older pups presented afterward to Day
22 or pregnancy-terminated females. This amplification of the hormonal
induction of maternal behavior by the special sensory qualities of neonates
may be important to the development of maternal behavior in parturient
rats. Rosenblatt suggested that shortly after parturition mother rats undergo
a transition from hormonally induced to nonhormonally maintained maternal
behavior (Rosenblatt, 1970; Rosenblatt et al., 1979). Stimuli associated
only with neonates may help to keep mothers interested in their young during
that transition.
This research was supported by National Science Foundation
Grant BN576-04316, awarded to M. B. Kristal. We thank Seymour Axelrod and
Harold I. Siegel for their valuable comments on an earlier version of this
paper, and Cindy Banas for the illustrations.
Reference Notes
1. Paul, L., Herrenkohl, L., Kupferschmidt, J., & Whitney, J. Conspecific young: The ability of the same stimulus complex to induce both cannibalism and maternal behavior. Paper presented at the Eastern Conference on Reproductive Behavior, Nags Head, North Carolina, May 1975.
2. Sachs, B. D. Behavior of maternal rats in the perinatal period. Paper presented at the meeting of the American Association for the Advancement of Science, Boston, December 1969.
3. Peters, L. C., & Kristal, M. B. Maintenance
and decline of the suppression of infanticide in mother rats. Manuscript
in preparation, 1983.
References
Dixon, W. J., & Massey, F. J. Introduction to statistical analysis. New York: McGraw-Hill, 1969.
Kristal, M. B. Placentophagia: A biohehavioral enigma (or De gustibus nondisputandum est). Neuroscience and Biobehavioral Reviews, 1980, 4, 141-150.
Kristal, M. B., & Graber, G. C. Placentophagia in nonpregnant rats: Influence of estrous cycle stage and birthplace. Physiology and Behavior, 1976, 17, 599-605.
Kristal, M. B., Whitney, J. F., & Peters, L. C. Placenta on pups' skin accelerates onset of maternal behavior in nonpregnant rats. Animal Behaviour, 1981, 29, 81-85.
Mayer, A. D., & Rosenblatt, J. S. Hormonal interaction with stimulus and situational factors in the initiation of maternal behavior in nonpregnant rats. Journal of Comparative and Physiological Psychology, 1980, 94, 1040-1059.
Moltz, H., Robbins, D., & Parks, M. Caesarean delivery and maternal behavior of primiparous and multiparous rats. Journal of Comparative and Physiological Psychology, 1966, 61, 455-460.
Paul, L., & Kupferschmidt, J. Killing of conspecific and mouse young by male rats. Journal of Comparative and Physiological Psychology, 1975, 88, 755-763.
Peters, L. C. The suppression of filicide in mother rats. Unpublished doctoral dissertation, State University of New York at Buffalo, 1980.
Rosenblatt, J. S. Views on the onset and maintenance of maternal behavior in the rat. In L. R. Aronson, E. Tobach, D. S. Lehrman, & J. S. Rosenblatt (Eds.), Development and evolution of behavior. San Francisco: Freeman, 1970.
Rosenblatt, J. S., & Siegel, H. I. Hysterectomy-induced maternal behavior during pregnancy in the rat. Journal of Comparative and Physiological Psychology, 1975, 89, 685-700.
Rosenblatt, J. S., Siegel, H. I., & Mayer, A. D. Progress in the study of maternal behavior in the rat: Hormonal, nonhormonal, sensory, and developmental aspects. In J. S. Rosenblatt, R. A. Hinde, C. Beer, & M.-C. Busnel (Eds.), Advances in the study of behavior (Vol. 10). New York: Academic Press, 1979.
Siegel, S. Nonparametric statistics for the behavioral sciences. New York: McGraw-Hill, 1956.
Slotnick, B. M., Carpenter, M. L., & Fusco, R. Initiation of maternal behavior in pregnant nulliparous rats. Hormones and Behavior, 1973, 4, 53-59.
Stern, J. M., & MacKinnon, D. A. Sensory regulation of maternal behavior in rats: Effects of pup age. Developmental Psychobiology, 1978, 11, 579-586.
Terkel, J., & Rosenblatt, J. S. Humoral factors under lying maternal behavior at parturition: Cross transfusion between freely moving rats. Journal of Comparative and Physiological Psychology, 1972, 80, 365-371.