Pergamon Press and Brain Research Publ.,1980.
Neophobia and Water Intake After Repeated
Pairings of Novel Flavors with Toxicosis1
MARK B. KRISTAL, MELISSA ANN STEUER, J. KEN NISHITA AND
LAWRENCE C. PETERS2
Department of Psychology, State University of New York at Buffalo, Amherst, NY 14226
Received 15 November 1979
Neophobia Taste aversion conditioning Lithium chloride
FLAVOR neophobia, the tendency for animals not to ingest novel-flavored substances, has been shown to be modifiable by the application of taste aversion conditioning procedures [2,3,4,5,11]. If a novel or relatively novel flavor is paired with toxicosis, enhanced neophobia can be observed either (a) during the toxicosis (illness-mediated neophobia) or (b) in association with the first novel flavor presented soon after aversion to the original novel flavor has been established (conditioned-aversion-mediated neophobia [2,4,5]). Since the studies involving the latter form of neophobia have usually involved testing for an aversion to the next one or two novel flavors a day or two after toxicosis, the extent to which the animal has learned that all novel flavors are dangerous, and therefore the generality of the neophobia, has not been probed. Although instances of increased finickiness or selectivity in conjunction with the induction of bait-shyness were reported years ago [9,10], we still do not know whether the animal's avoidance is because all novel substances are considered dangerous (generalized neophobia), whether only novel substances presented soon after conditioning are considered dangerous, or whether only substances similar to the conditioned stimulus are considered dangerous (simple stimulus generalization).
We decided to test for the formation of a generalized conditioned neophobia by administering a series of novel flavors to naive rats at 4-day intervals, and following each presentation with an injection of a toxicosis-inducing drug. The formation of an aversion to the presented flavors was not assessed during the series of presentations, since such a trial (a) would introduce a non-novel flavor into the series, and (b) would actually constitute an extinction trial. The object, therefore, was to observe how many presentations of novel flavor with toxicosis were required to cause the rats to stop ingesting novel flavors when they were presented. After the series of conditioning trials ended, the maintenance of an aversion to the first novel flavor presented was assessed, after which still another novel flavor was presented to test for the maintenance of the neophobia after an extinction trial.
The rats in the experiment were all placed on, and stabilized to a 23½-hr water-deprivation schedule. On flavor-presentation days only novel-flavored solution would be available for the first 15 min of the drinking period, and only plain water would be available for the second 15 min. Therefore, we were able to test for a second phenomenon: whether suppressed drinking of the novel flavored solution and discrimination between the flavored solution and plain water (see [1,6,7] for review) would lead to increased drinking of plain water in order to maintain daily water intake level.
METHOD
Animals
Thirty-five male, Long-Evans rats (Charles River Breeding Laboratories) approximately four months old, and weighing 250-300 g were used in the experiment. The rats were housed individually in 24 x 19 x 18-cm hanging wire-mesh cages fitted with food hoppers (containing Charles River Rat/Mouse/Hamster Formula 3000) and with water bottles (inverted plastic l00-ml graduate cylinders with stoppers and drinking tubes). The rats were maintained on a 14:10 day:night cycle, with the day phase beginning at 0600 hr (EST).
Procedure
Groups. All 35 rats were trained for two weeks to drink water on a 23½-hr water deprivation schedule, after which they were assigned either to the experimental group (n = 15) or to one of two control groups (n = 10 each). Rats were assigned to the three groups such that the groups were matched for baseline water intake just prior to the start of the experiment (mean ± SEM ml for each group: 20.44 ± 0.54, 20.20 ± 0.58, and 20.10 ± 0.50, over the last three days of baseline stabilization). Each of the experimental rats (TAC Group -- taste aversion conditioned) received an injection of a toxicosis-inducing drug (20 ml/kg body weight of 0.15M lithium chloride, administered IP [8]) 15 min after each 15-min presentation of the 5 novel flavors in the series. One control group (S Group -- saline injected) was used to assess the effect of injections that followed novel-flavor presentations. Injections for S Group rats consisted of 20 ml/kg body weight of physiological saline, injected IP. The second control group (NC Group -- noncontingent LiCl injected) was used to assess the effect of repeated toxicosis. For NC Group rats, a 20 ml/kg body weight, IP injection of 0.15M lithium chloride (LiCl) was administered 2 days after the presentation of a novel flavor. Therefore, the rats of NC Group were getting the same number of poisonings as those of the TAC Group, but the bouts of illness were not contingent upon novel-flavor ingestion.
Novel-flavored solutions. The five novel-flavored drinking solutions were presented in five counterbalanced orders of presentation. In the TAC Group there were three rats on each of the five orders of presentation, and two on each order in each of the control groups. The concentrations used for the novel-flavored solutions were those which had been determined to lead to approximately equal subsequent water intakes when presented to a separate pilot group on the same water-deprivation and flavor-presentation schedules as the test rats. The novel flavors used were: beef bouillon (MBT®, beef-flavored, in packets) 10% w/v; grape juice (Welch's® Grape Juice, unsweetened), 50% v/v; lime drink (Kool-Aid®, lime-flavored, unsweetened), 2.5% w/v; powdered milk (Carnation® Instant Nonfat Dry Milk), 10% w/v; and chocolate drink (Faygo® Chocolate Creme flavored soda, artificially sweetened, decarbonated), 50% v/v. The five flavors represent different levels of palatability. Since the flavors were presented in counterbalanced fashion, all five flavors were presented on the first day (each flavor to 7 of the 35 rats). These first-day intakes were used as measures of baseline attractiveness of each of the flavors to as yet untreated rats. Therefore subsequent intake of a particular novel-flavor was calculated as a percentage of the baseline intake for that flavor. The baseline intake (mean ± SEM ml) for each of the five novel flavors was: beef bouillon, 20.0 ± 2.43; grape juice, 5.29 ± 0.90; lime drink, 6.29 ± 0.93; powdered milk, 15.57 ± 2.34; chocolate drink, 13.29 ± 1.87.
A sixth novel flavor was presented after the series of five flavor presentations. All rats received the same sixth novel flavor, orange drink (Wylers® Orangeade, sugar added, 25% w/v). Since computing of a baseline was not possible because all rats received it only on the last presentation, absolute intake values of this flavor (in ml) were used.
Presentations. On Days 1, 5, 9, 13 and 17 of the experiment, at about 1600 hrs, each rat in the TAC Group received 15 min of access to one of the novel flavors, followed immediately by 15 min of access to tap water (all drinking solutions were presented at room temperature in thoroughly cleaned graduate cylinders). At the end of the 15 min of access to water, the water was removed and each rat received the LiCl injection. Rats in S Group were treated identically, except that the injection contained physiological saline rather than LiCl solution. For the NC Group, the LiCl injections were administered after the half hour of water drinking that occurred two days after novel-flavor presentation (i.e., LiCl injections only on Days 3, 7, 11, 15, and 19). On Day 24 of the experiment (7 days after the fifth novel-flavor presentation) all rats were presented with the novel-flavored solution they had been given on Day 1. Intake of that substance (Presentation VI) was used to measure the persistence of the aversion which had presumably been produced by the first LiCl injection, but which had not been tested for previously. No injection was administered after Presentation VI. Seven days later (Day 31) all the rats were presented with yet another novel flavor (Presentation VII -- orange drink) to assess the effects of time and an intervening extinction trial (Presentation VI) on the neophobia produced during the initial series of five flavors.
RESULTS
Intake of Novel-Flavored Solutions
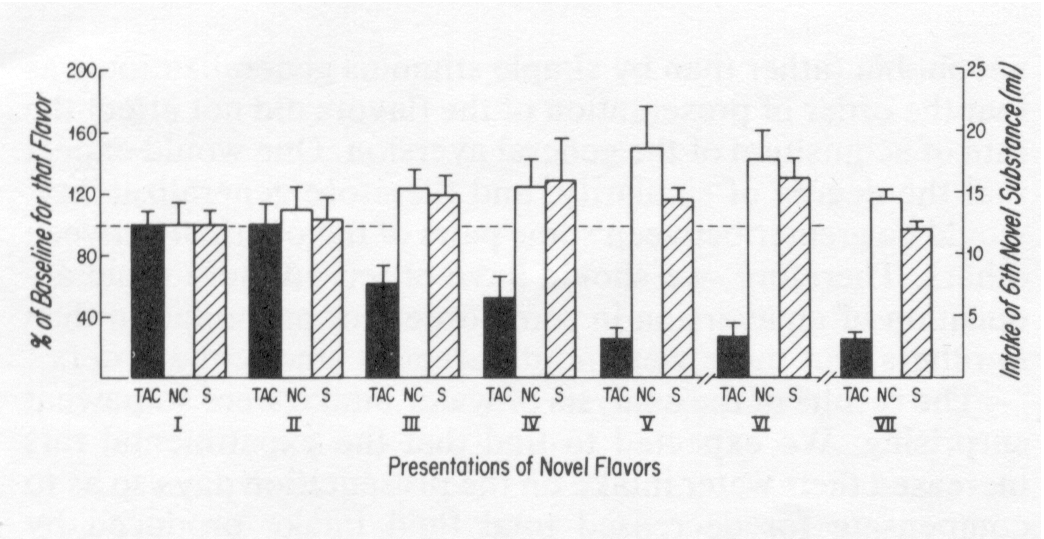
Figure 1 contains the group values for the percentage of baseline intake (per novel flavor) on each of the presentations. As mentioned earlier, the mean intake on Presentation I for all rats receiving that particular flavor (7 rats/flavor) was taken as the baseline intake for that flavor. Subsequent intakes were computed as a percentage of the baseline for that flavor. Figure 1 shows that by Presentation III, the group receiving a contingent LiCl injection after each presentation (TAC Group) was manifesting a statistically significant suppression of novel-fluid intake. The intake percentages for the three groups on Presentation III were analyzed with an ANOVA followed by a Scheffé multiple contrast test. The TAC Group's intake of novel-flavored fluid was significantly less than the intakes of the two control groups, which did not differ from each other, ANOVA: F(2,32) = 9.02, p < 0.001; Scheffé: TAC vs S, F(2,32) = 11.10, p < 0.01; TAC vs NC, F(2,32) = 13.60, p < 0.01; NC vs S, F(2,32) < 1.0.
A 2-way ANOVA (for repeated measures on one factor, using a Greenhouse-Geisser conservative F-test for the Within Subjects analysis [12]) was used to compare the effect of number of presentations of novel flavors with the effect of the order of presentations of the novel flavors, on novel-flavored solution intake. As in the previous analysis, the effect of number of presentations was significant for the TAC Group but not for the others. The effect of order of presentation was not significant, F(4,30) = 2.76, p > 0.01, nor was the effect of the interaction of order of presentation with number of presentations, F(4,30, for conservative test) = 2.50, p > 0.01.
Maintenance of the aversion assumed to have been formed in the TAC Group rats after the first pairing of novel flavor with toxicosis was ascertained by comparing the intake of the novel flavor available during Presentation I with intake of that same flavor during Presentation VI. The results of the analysis indicated a significantly lower intake on Presentation VI, (t-test for paired observations: t(14) = 7.42, p < 0.005), which suggests that the aversion formed at Presentation I was still in effect after four subsequent pairings with other novel flavors.
FIG. 1. Effect of number of pairings of novel-flavored solutions with toxicosis on intake of novel-flavored solutions. Presentations (15 min of access to novel-flavored solution followed by 15 min of access to water) I through V were followed immediately by an injection of LiCl (TAC Group, n = 15), immediately by an injection of saline (S Group, n = 10) or after two days by an injection of LiCl (NC Group, n = 10). Presentation VI consisted of re-exposure to the flavor presented in Presentation I. Presentation VII consisted of the same entirely novel substance for all rats. Presentations VI and VII were not followed by injections. The results (except for those of Presentation VII) are collapsed over 5 counterbalanced orders of presentation of 5 novel flavors. Note that although the results of Presentations I through VI are in mean (± SEM) % of baseline intake, the results of Presentation VII are in mean (± SEM) ml intake.The intake of novel-flavored fluid in Presentation VII (orange drink) was compared among the groups using a 1-way ANOVA followed by a Scheffé test. The intake values of the TAC Group were significantly lower than those of the two control groups, which did not differ from each other, ANOVA: F(2,32) = 85.20, p < 0.001; Scheffé: TAC vs S, F(2,32) = 86.40, p < 0.001; TAC vs NC, F(2,32) = 143.10, p < 0.001; NC vs S, F(2,32) = 6.04, p > 0.01. Therefore, after a presentation without aversive consequences (Presentation VI) and two weeks after the most recent toxicosis, the experimental rats still manifested a strong reluctance to drink a novel-flavored solution.
Water Intake
Figure 2 depicts the daily mean total fluid intake per group (in ml) over the course of the experiment. Although all groups showed a slight depression in total fluid intake on the day after injection for the first couple of trials, the effect was insignificant. Eventually though, the rats in the TAC Group began to show a dramatically reduced total fluid intake on the day of the presentation, whereas the other groups did not. Reduced intake on the day of presentation in the TAC Group indicated that either the TAC Group rats were holding water intake constant and decreasing intake only of the novel-flavored solution, or that they were reducing the in take of both novel-flavored solution and of plain water. Figure 3 contains the intake only of water (second 15 min of drinking period) on presentation days. The values are expressed in terms of the percentage of baseline (Presentation I) intake. Clearly, intake of plain water is reduced rather than maintained. A 1-way ANOVA and Scheffé test comparing the water intake values on Presentation VI for the three groups indicated that the TAC Group intake was significantly lower than those of the two control groups, which did not differ from each other, ANOVA: F(2,32) = 8.44, p <0.001; Scheffé: TAC vs S, F(2,32) = 7.53, p < 0.05; TAC vs NC, F(2,32) = 14.82, p <0.01; NC vs S, F(2,32) = 1.04, p > 0.05. The difference between the TAC Group and the controls had not yet reached significance on Presentation V.
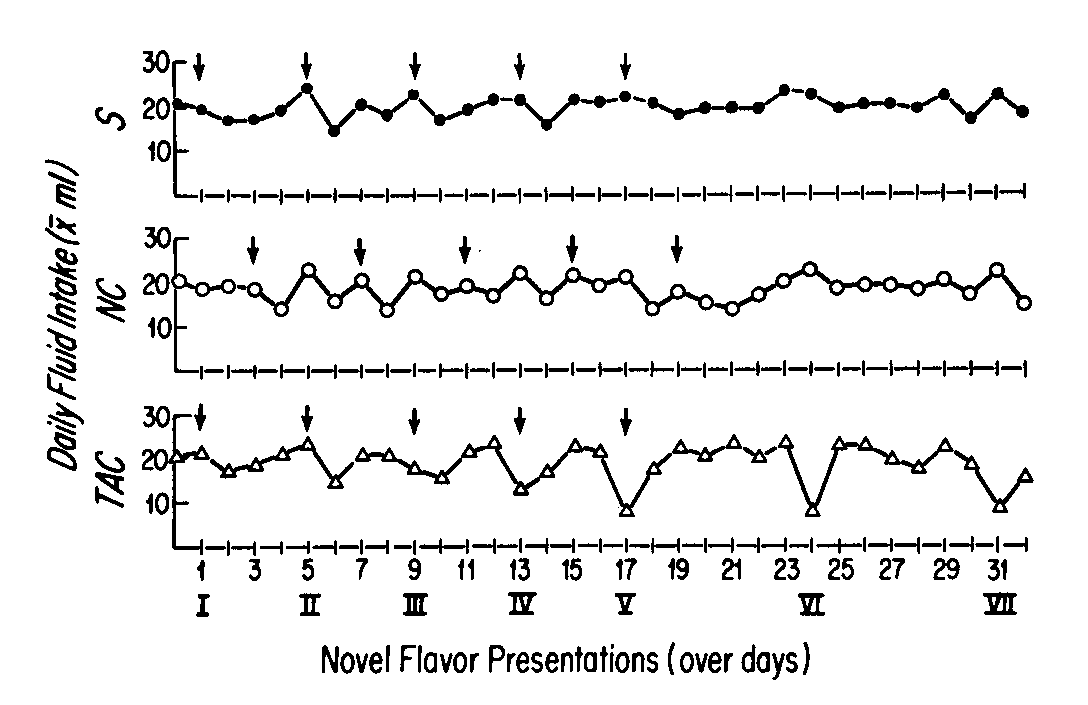
FIG. 2. Total fluid intake (mean ml) per 30-min drinking period per day of the experiment. Roman numerals represent days on which novel-flavored drinking solutions were presented instead of plain water for the first 15 min of the drinking period. Arrows indicate days on which injections of LiCl (TAC and NC Groups) or saline (S Group) were administered immediately after the drinking period. Standard errors of the mean were too small to be represented accurately in the figure. They ranged from 0.64 ml (TAC Group on Day 29) to 2.80 ml (NC Group on Day 1), with 90% falling between 0.70 and 1.60 ml.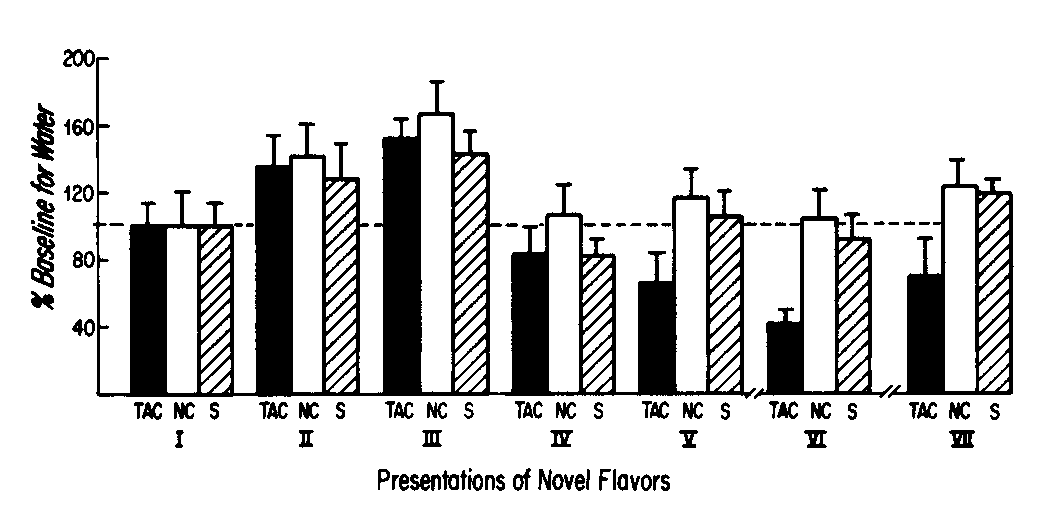
FIG. 3. Water intake on the days of presentation of novel-flavored solutions. Group intake of plain water, which was available only for 15 min after 15 min of access to novel-flavored drinking solutions, is expressed as a percentage of baseline intake for that group (mean ± SEM %). Group treatment and sizes are those described for Fig. 1. In contrast to Fig. 1, however, the units of measurement for the results of Presentation VII, in the present figure, are the same as those for the other six presentations.
DISCUSSION
Through the use of taste aversion conditioning procedures we have attempted to demonstrate that rats can be taught a generalized neophobia. Formerly, rats were only required to express conditioned neophobia in studies testing for the response toward one or two novel flavors presented soon after an aversion was established. It was not known, therefore, whether the rats had learned that novel substances, per se, were to be avoided, or whether the first, the most similar, or the next day's novel substances were to be avoided. Furthermore, the previous studies were confounded by testing for the presence of an aversion (thereby introducing extinction trials) before testing for the acquired neophobia. In the present study we have sought to eliminate these problems by presenting a number of different flavors paired with toxicosis, and observing when in the series of presentations the rats begin to suppress their intake of novel-flavored solutions.
Since the order of presentation, the temporal pattern (by using a 7-day rather than 4-day interval between Presentations V and VI and between VI and VII), the injection procedure, and the effect of repeated poisonings were all controlled for, we think the argument that the rats were responding to the novelty dimension as a cue, is convincing. It is not possible, of course, to rule out stimulus generalization based on some perceived similarity, other than the novelty dimension, among the flavors chosen. However, although all the flavors we proffered had to be palatable, we determined empirically that they were of different degrees of palatability, and estimated subjectively that there were major qualitative differences among the flavors. This suggested to us that the rats should have readily distinguished among the flavors. An additional piece of evidence suggesting that the behavior was mediated by the formation of a general neophobia rather than by simple stimulus generalization was that the order of presentation of the flavors did not affect the rate of acquisition of the general aversion. One would expect that the degree of similarity, and therefore generalizability, would be greater between some pairs of flavors than between others. Therefore, we should have observed more rapid acquisition of an aversion in some orders of presentation than in others, but there were no differences among the orders.
The results of the analysis of water intake were somewhat surprising. We expected to find that the experimental rats increased their water intake on the presentation days so as to compensate for decreased total fluid intake produced by avoidance of the novel-flavored solution. Rather than make a fine distinction between the "dangerous" novel flavor, and "safe" tapwater, as we would have expected from the literature, the rats apparently resolved the problem by sup pressing all drinking on that day. It should be noted that a simple conditioned aversion to water was not what was established. The rats in the NC Group received an LiCl injection immediately after 30 min of access to plain water, but neither formed an aversion to plain water, nor formed a temporal suppression pattern based on a 4-day injection schedule. We cannot say, on the basis of this study, whether the water-intake suppression was due to learned suppression of drinking, to an unwillingness or inability to discriminate, or to a general decrement in consummatory responding. We can say, though, that under certain circumstances, the water-deprived rat finds it preferable not to drink than to decide what to drink.
1Supported by NSF Grant BNS76-04316 awarded
to M.B.K. We wish to thank Bennett G. Galef, Jr., Kenneth J. Levy, and
Seymour Axelrod for their comments on an earlier version of this manuscript.
Our thanks to J. F. Whitney for his help in collecting data.
2 Now at the Institute of Animal Behavior, Rutgers University.