![]() Top
Top
![]() Abstract
Abstract
![]() Introduction
Introduction
![]() Methods and Materials
Methods and Materials
![]() Results
Results
![]() Discussion
Discussion
![]() References/Acknowldgements
References/Acknowldgements
|
|
|---|
Theodore H. Bullock(1), Sacit Karamürsel(2), Jerzy Z. Achimowicz, Michael C. McClune and Canan Baar-Eroglu(3)
Neurobiology Unit, Scripps Institution of Oceanography
and
Department of Neurosciences, School of Medicine,
University of California, San Diego
La Jolla, California 92093-0201 U.S.A.
Running title: Human VEP and OSP
Correspondence to: T.H. Bullock
Department of Neurosciences
University of California, San Diego
9500 Gilman Dr. DEPT 0201
La Jolla, CA 92093-0201, U.S.A.
Ph: 619/534-3636; Fax: 619/534-3919;
E-mail: tbullock@ucsd.edu
|
|
Abstract |
|
|
|---|
Visual evoked potentials (VEP) and omitted stimulus potentials (OSPs) are reexamined in scalp recordings from 19 healthy subjects. The principal finding is a distinction in form, latency and properties between OSPs in the conditioning stimulus range <2 Hz, used in previous human studies, and those in the range >5 Hz, used in previous studies of selected elasmobranchs, teleost fish and reptiles. We cannot find OSPs between 2 and 5 Hz. The high frequency ("fast,"ca. 6 to >40 Hz) and the low frequency ("slow," ca. 0.3-1.6 Hz) OSPs have different forms and latencies but both tend to a constant latency after the omission, over their frequency ranges, suggesting a temporally specific expectation. Fast OSPs (typically N120, P170-230 and later components including induced rhythms at 10-13 Hz) resemble an OFF effect, and require fixation but not attention to the ISI. Slow OSPs (usually P500-1100) require attention but not fixation; they are multimodal, unlike the fast OSPs. Based on cited data from fish and reptiles, fast OSPs probably arise in the retina, to be modified at each subsequent level. We have no evidence on the origin of slow OSPs. In both ranges not only large, diffuse flashes; but weak, virtual point sources (colored LEDs) meters away suffice. They are difficult to habituate. Both require very short conditioning periods.
The transition from the single, rested VEP to the steady state response (SSR) at different frequencies is described. Around 8-15 Hz in most subjects larger SSRs suggest a resonance. Alternation between large and small SSR amplitude occurs around 4 Hz in some subjects and conditions of attention, and correlates with an illusion that the flash frequency is two Hz or is irregular. Jitter of the conditioning intervals greatly reduces the slow OSP but only slightly affects the fast OSP. Differences between scalp loci are described.
Key words: Cognitive potential; Event-related potential; Steady state response; Expectation; Attention
Abbreviations: EEG, electroencephalogram; EP, evoked potential; ERP, event-related potential; IR, induced rhythm; ISI, interstimulus interval; LFF, lowest frequency for complete fusion in averages; OSP, omitted stimulus potential; P75, positive peak at 75 ms; SSR, steady state response; UFF, upper frequency for 1:1 following in single sweeps.
|
|
Introduction |
|
|
|---|
Event-related potentials (ERPs) to missing stimuli have been known in human subjects for many years (Rusinov 1960; Barlow et al. 1965; Sutton et al. 1965, 1967; Klinke et al. 1968; Picton et al. 1974; Renault and Lesevre 1977, 1978; Squires et al. 1975, 1977; Kerkhof 1978; Courchesne 1979; Ritter et al. 1979; Ruchkin and Sutton 1979a, b; McCallum 1980; Ford and Hillyard 1981; Takasaka 1985; Hillyard and Picton 1987; Alain et al. 1989; Mangun et al. 1993). However, their dependence upon the interstimulus interval (ISI) of the series prior to the omitted stimulus, as well as upon the duration and regularity of the series have received little attention. These dynamic properties became important when it was discovered that ERPs to omitted stimuli are prominent in nonmammalian vertebrates. They are already well developed in the midbrain tectum and even in the retina after conditioning series of light flashes in rays and turtles (Bullock et al. 1990; Prechtl and Bullock 1992, 1994) as well as in primary sensory nuclei of the medulla after physiological electrosensory stimuli in rays (Bullock et al. 1993). In these studies the most effective conditioning frequencies were found to be in a range (>2 Hz) not heretofore explored in human subjects. The latency of the OSP in these animals is much more precisely constant after the due-time of the omission, over a range of conditioning ISIs, than has been noted in humans, suggestive of a temporally specific expectation. The effect of irregularity of ISI (hereafter "jitter") is minimal in these species of fish and reptiles in their effective frequency range for OSPs, whereas the limited information on humans (Takasaka 1985), indicates that jittered stimulus series give no average OSP. The aims of the present study are to examine the neglected range >2 Hz in normal human subjects, to measure the latency of the OSP after different conditioning frequencies, to report a surprisingly short minimum conditioning series and a more careful test of the effect of irregularity of ISIs. Relevant to these questions also is better information on the transition of visual evoked potentials (VEPs) from the rested response after a single flash to the steady state response (SSR) at different ISIs. We report in addition on amplitude alternation of successive VEPs under certain conditions, including attention, on the effects of binocular stimulation, cross-modal stimulation, and differences in dependence on attention in the "high" (>2 Hz) and "low" (<2 Hz) frequency ranges. The occurrence under some conditions of induced rhythms, fluctuations in sensitivity and regional differences in response are noted.
|
|
Materials and Methods |
|
|
|---|
Subjects
Under protocols from the human subjects committees of the Universities of California and of Lübeck, 19 normal volunteers, clinically normal, high functioning adults, aged 30-75 (4 female), mostly scientists familiar with the project, were included in this study. No differences in results appeared to be correlated with age or gender. During the recording session, subjects sat in a chair in a dimly illuminated room, in a relaxed position. They were usually asked not to move or speak, or to blink too much, and to look at a fixation point on the flash probe. Subjects were instructed to try to predict the time of the next flash or, in the case of low frequency stimulation with single omissions, to count the omissions. In the case of low frequency stimulation with jittered ISIs, they were instructed to try to predict the time of the next flash. In the case of high frequency (>5 Hz) stimulation, usually given in short trains to permit seeing the whole OSP without subsequent VEPs, they were given no instruction. Above ca. 4 Hz, subjects report it soon becomes impossible to keep track of each flash or its absence. Short rest periods were provided between recording sessions. The number of recording sessions in a day together amounted to 2.5-3.5 hours.
Recording
EEG data were taken via Ag-AgCl disk electrodes or gold electrodes (Grass). The active electrodes were placed according to the international 10/20 system at Cz, Fz, Oz, O1, O2, P3, and P4, or at some of them. The reference electrode was on the left ear and grounded electrodes on both ears. EEG signals were amplified via Grass amplifiers with 60 Hz notch filters and the output sent to steep analog bandpass filters (Rockland, model 452) set variously, from 0.3-100 Hz to 0.1-50 Hz. EEG data were collected in a PDP 11/73 computer which also controlled the stimuli. The EEG data were monitored by oscilloscopes and a Nicolet 1170 computer during the recording sessions. Off-line analysis was performed on the PDP 11/73.
Stimulation
Visual stimuli (8 µs pulses of bright flashes) were generated with a Grass PS 2 photo-stimulator or a glow modulator driven by the PDP 11/73 computer and/or a PulseMaster A300 digital stimulator. Flash intensity with the Grass strobe light was #1 or #2 for all paradigms and the flash unit was covered with paper and placed 0.5 -1 m from the subject. Small green LEDs were also used for visual stimulation. Auditory stimuli were given as binaural clicks via a stereo headphone.
Stimulus Paradigms
Single flash VEPs were taken with at least 4 s intervals (ISI) and the effect of the stimulus intensity and the ambient illumination were investigated in a few subjects. The SSR and the OSPs to visual stimulation were recorded at 0.3, 0.5, 1, 2, 4, 5, 6.7, 10, 15, 20, 30 and 40 Hz flash frequencies. Two stimulus regimes were used. In the train paradigm 2-30 s trains (conditioning periods) were used after 2-30 s rest periods. In the continuous series paradigm one stimulus was omitted every 5-10 flashes, pseudorandomly. In a variant of the continuous series, the OSP to omission of every third stimulus was compared with those after omission of the 4th, 5th or 6th stimulus in a regular series.
Cross-modality effects were found in the equivalence, for certain frequencies, of a terminal stimulus of a different modality (click ending a flash train or flash ending a click train).
To test for the effects of irregularity of intervals the ISIs were jittered in a low and a high percentage of the mean, with the condition that the last interval was equal to the mean.
Experiments were done in two different periods in Lübeck and an extended period in La Jolla, with different setups and overlapping personnel. Most of the techniques and procedures were quite standard or differed in ways that proved quite insignificant, according to the results, for example placement of electrodes, filter settings and methods of rejecting sweeps. No doubt, however, there were unintentional differences that might have affected results, for example in instructions to subjects and state of the subjects. Our judgement is that these were no greater or more systematic between laboratories than within each laboratory. Uncontrolled differences between runs or subjects or months surely occurred in each locale. The extremes of the variance of results under the same stimulus regime could occasionally be found in the same subject, some months apart, but successive sessions on the same day or different days, up to months apart gave quite the same results in each subject in >95% of such comparisons. In this basically descriptive sampling of human responses we have not undertaken a quantitative study of the variance of selected measures but report the ranges of most measures qualitatively.
Analysis
The average EP, SSR and OSP responses were obtained from up to 75 to 100 single sweeps. Selective averaging was used to eliminate movement and eye blink artifacts, i.e. sweeps judged by eye to contain such artifacts were rejected. Steep digital lowpass filters removed high frequency artifacts and EEG components in some experiments.
|
|
Results |
|
|
|---|
The principal surprising result is the distinction between OSPs in high and low frequency conditioning stimulus ranges (B2, below). Before detailing this finding, we will mention some of the stimulus parameters that can influence the form, amplitude or latency of OSPs. This will involve some new and unexpected dynamic aspects of human scalp VEPs which are the responses to the background train of flashes that must be distinguished from those attributable to omission.
A. Effects of temporal parameters on OSPs and preceding VEPs
Prolonged stimulation does not substantially influence the measures reported here; they do not habituate or fatigue. In repeated records during an hour or more of nearly continuous stimulation it is usual to see no changes in the VEPs in either the low flash rate (<2 Hz) or high rate (>2 Hz) range; each frequency reaches a steady state response after a short transition period and maintains this SSR.. This observation is not from examining single sweeps but from comparing averages of 50 sweeps; we cannot say what changes may have occurred within that time. The OSPs to low and high rate stimulation likewise do not change with time unless the subject loses attention, which has to be of a different sort for the two ranges (see C., below).
OSPs to the omission of a single stimulus during a sustained train are similar in morphology to those after the end of a train. They may look more complicated in the former paradigm due to the superimposition of the next VEPs and their transition to an SSR in the high rate range (Fig. 1). At slow rates OSPs to the omission of a single stimulus and to the end of a train are quite the same in form.
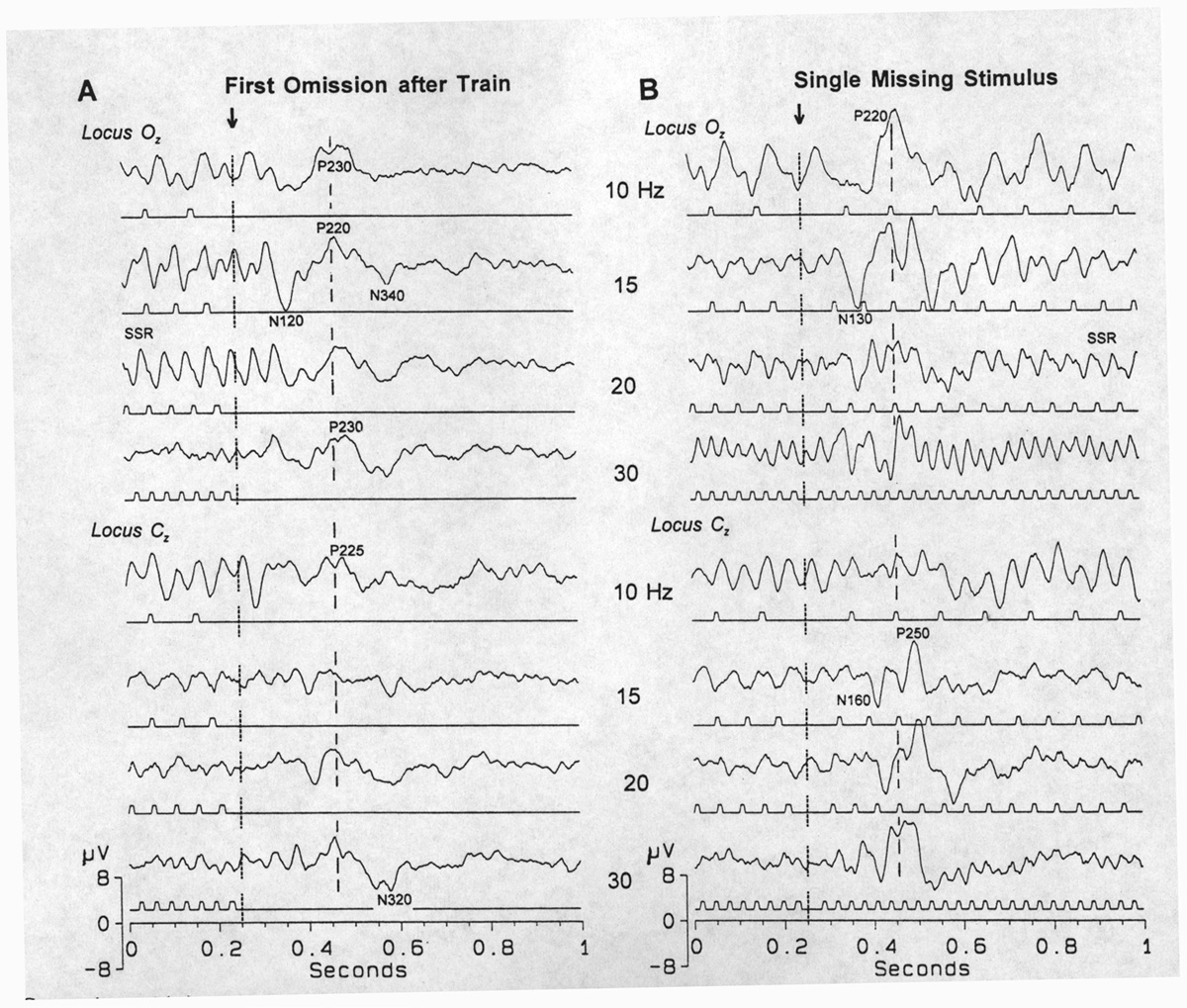 |
Fig. 1. Comparison of (A) the end of a train and (B) the omission of one stimulus in a high frequency range of stimuli (10-30 Hz), in two loci (upper, Oz; lower, Cz), aligned by the due-time of the first omitted stimulus. The main peaks of the OSP - some are marked with their polarity and latency after the time of omission - are almost the same in the two paradigms but look more complicated in B due to the superimposition of the slightly rested VEP and the SSR to stimuli after the omission. Averages of 100. |
A1. Train duration, stimulus number and interval between
trains.
Each of these parameters can exerts some influence on the shape
and amplitude of VEPs and OSPs but the effects are not large.
In the high frequency range, tests from 5-40 Hz show
that a 2 s conditioning period gives OSPs as large as does a 30
s period and the same is true after 2 s and 30 s resting periods
following either the long or the short conditioning. We have not
studied the beginning of the train at these frequencies in human
scalp recordings to see the transition from the first, rested
VEP to the SSR but this is reported for the elasmobranch in Karamürsel
and Bullock (1994), where it is shown that much less than 2 s
is required for an OSP and only a very few ISIs..
In the low frequency range, tests from 0.3-2 Hz show that the VEP comes to its final form and amplitude after the second stimulus. Amplitude, latency and shapes of the 3rd, 4th and 5th VEPs are exactly the same as the second one, if we examine averages of trains. This was true in each of the six subjects examined for this point but we expect to find exceptions since the VEP is so variable in form and amplitude among subjects. We did not study this point quantitatively or at many scalp loci. In the low frequency range OSPs also show a low minimum requirement for preceding ISIs; the amplitude, latency and shape of the OSPs to the omission of the 3rd, 4th, 5th and later stimuli look the same. At 0.3, 0.5 and 1 Hz, two stimuli are enough, i.e. one ISI, to elicit an end of train wave that looks just like a true OSP (Fig. 4A). It is important, however, to point out that these comparisons are made on averages, so that the system has seen a patterned series of stimuli and omissions. According to the conclusions from similar experiments on elasmobranch fish (Karamürsel and Bullock 1994), we can expect that, if the SSR has developed and stabilized, so that the next sequence can be predicted, the conditioning period of that duration is long enough to show a good OSP. This implies that the response to the omission of every third stimulus (= omission of 3rd, 6th, 9th, etc.) is generated by a mechanism similar to that for omissions of every fourth or fifth or sixth, in this low frequency range.
A2. Effects of stimulus frequency on VEP and OSP.
No OSP has been observed in any of our subjects to 2-5 Hz flash
stimulation; the VEPs are not followed by an OFF response. Good
OSPs are obtained below 2 Hz (1, 0.5 and 0.3 Hz), and above 5
Hz (6.7, 8, .., 40 Hz). A transition zone between 1 and 6.7 Hz
separates OSPs of different characteristics for slow and fast
stimulation rates. Accordingly, we describe two types of OSP in
humans (Figs. 1, 3, 4). Those following fast and slow conditioning
stimulus trains we will call fast and slow OSPs.
A2a. High stimulus rate (>2 Hz).
The amplitude of the OSP becomes greater with higher conditioning
rates in the range of 5-15 Hz and then stays unchanged or in some
subjects declines. The SSR becomes smaller with higher frequency
and disappears above 20 Hz in most subjects. The lowest frequency
at which the SSR disappears, even in an average, is the LFF or
lowermost fusion frequency by the evoked potential criterion.
It is not the same as the subjective flicker fusion frequency,
nor is it a fixed value but depends on intensity, ambient light,
peripheral vs central vision and other factors. It can be as low
as 20 Hz - in the same range as we have reported for many of the
slower fish species, whereas other fish species are much higher
(Bullock et al. 1990).
Around 8-15 Hz, in some loci in most of the subjects, the SSR is much larger than expected from the amplitude at lower rates, suggesting a resonance (Fig.1A, 15Hz; Fig. 3, Oz, 10-15 Hz).
The latency of the OSP after the next due-time is quite constant in the stimulus frequency range of 10-15 to 40 Hz, in the same subject. Since below 10 Hz, OSPs are small, longer, more complicated, superimposed on the SSR and sometimes with oscillatory components, it is difficult to determine their latencies with accuracy. Variability is considerable between subjects but it is not amenable to quantitative comparison; like the VEP, the form, initial polarity and number of peaks as well as amplitude vary between subjects in scalp recordings, possibly due to differences in the geometry of the striate convolutions. In most subjects, the OSP appears as an N120-P170 and P200-P250. Later components are often seen, at least to 500-700 ms, but are even more variable.
Alternation of successive responses during an SSR between large and small VEPamplitude ("1 in 2") is observed in some subjects (in six out of nine where it was looked for; four of these showed it well), at certain stimulus rates, commonly at 4 Hz (Fig. 2). The SSR follows the flashes 1:1 in most
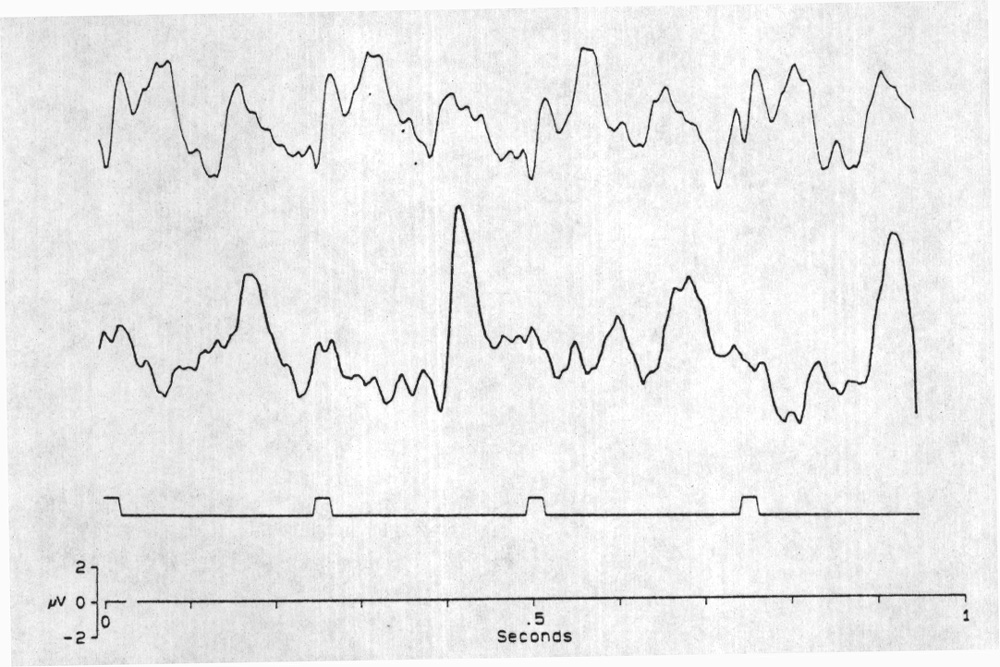 |
Fig. 2. Alternation in the amplitude of successive VEPs and a correlated illusion. Upper trace is the SSR to 4 Hz stimulation, under conditions that cause 1:1 following at constant amplitude - moderate intensity (Grass Photostimulator, intensity #1), diffused flash, ca. 1 m from the subject. Vertex derivation. Each trace the average of 128 sweeps. Second trace, 4 Hz but a special state of attention changes the SSR following to 1 in 2 (alternately smaller and larger VEPs); this must be quite regular to be preserved in such a long average. At this time, the subject reports that the flash rate appears to be 2 Hz. Often when the apparent rate is higher than 3 Hz, a certain kind of attention causes the illusion that it is irregular. Sometimes the difference between alternate amplitudes is small and it may be evident only in one or some of the leads. Possibly the regularity is imperfect and averaging smears the alternation. A correlate is that, subjectively, it is difficult to hold the illusion for long. Averages of 74 sweeps; amplifier bandpass 0.3-55 Hz.The stimulus markers are artificially prolonged for visibility; the time of the brief flash is the onset of the mark. |
subjects, even at high frequencies, up to nearly 20 Hz. Every other VEP is larger under special conditions, however, including no eye blinking during the whole recording session, careful fixation to one point on the flash probe held within 10-15 cm from the face and possibly other, unrecognized conditions. In that state there is an illusion; subjects report that the 4 Hz flash frequency is 2 Hz. This seems important, because it is a strong indication of the effect of attention on VEPs. Alternations in SSR amplitude are not seen often, as they are in rays (Karamürsel and Bullock 1994), perhaps because scalp recording averages over too much cortex or because it requires averaging over many trials and alternation is lost in averaging if it does not always begin with the same phase or is not perfectly consistent during the sampling period. We usually need to average at least 30 sweeps of one second each to see a clear response, and hence to maintain the same state of close attention unchanged for at least 30 s. Figure 3 shows some 1 in 2 following at 4 Hz in locus Pz.
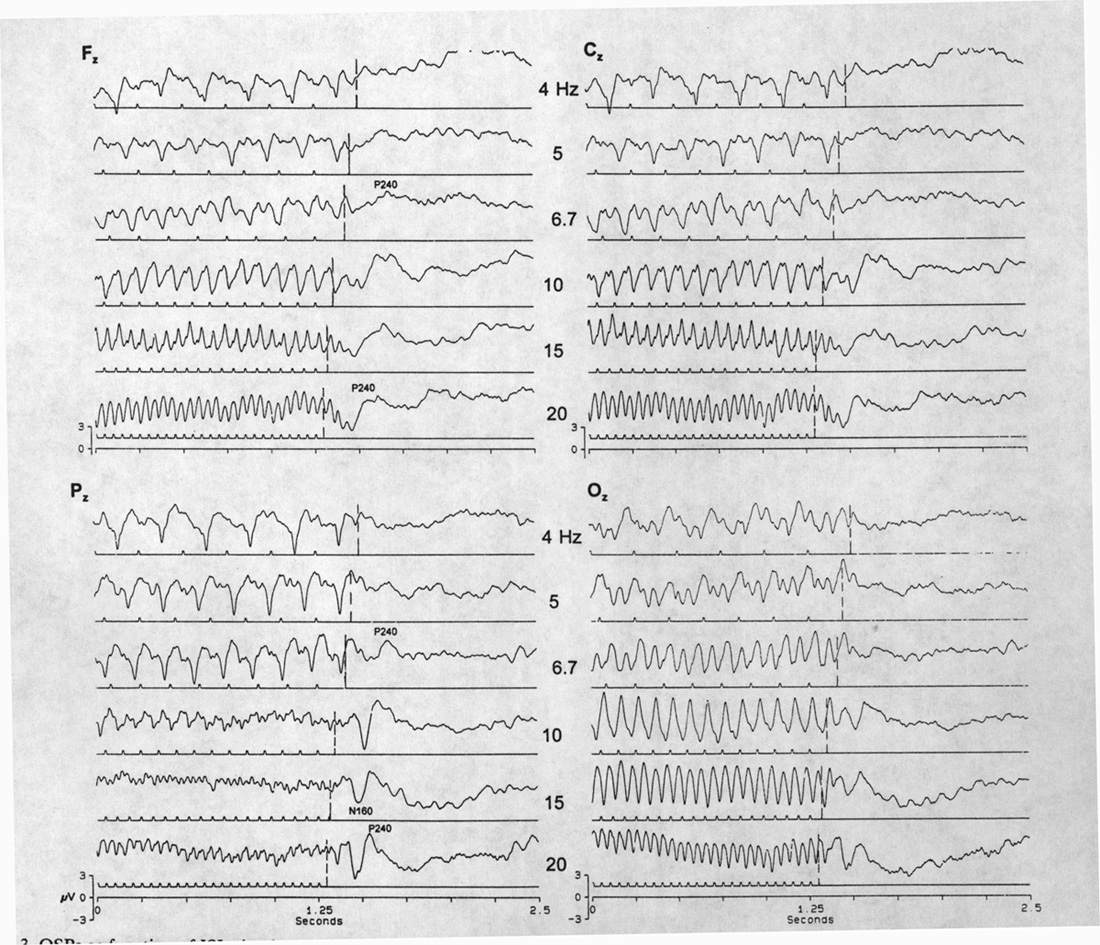 |
Fig. 3. OSPs as function of ISI; simultaneous recordings from four sites: midline frontal, central, parietal and occipital. Traces are aligned by the last flash. Due-time of first omitted stimulus is shown by vertical dashed lines. Note that the best OSP is in Pz, mostly N170, P230 after the first missing stimulus due-time at high frequencies (10, 15 and 20 Hz). The P230 alone appears first at 5 Hz (Pz). The early components of the OSP responses are smaller in Oz whereas the SSR waves are larger. The late components of the OSPs are much the same in the four loci. Averages of 75 sweeps. |
OSP alternation is not usually observed in averaged records at higher rates; this does not mean it has not occurred, for the reasons just stated. At 20, 30 and 40 Hz, however, the SSR sometimes has a repeating pattern that suggests a larger VEP about every 7th flash.
A2b. Low stimulus rate (<2 Hz).
VEP amplitude gets smaller with higher stimulus rates. A
late component (N250-350) which is conspicuous in the rested VEP
(0.1 or 0.25 Hz) is more sensitive than earlier components. This
late negative wave is always reduced or absent during the conditioning
period in experiments on OSPs at the low rates of 0.3 - 1 Hz.
We have not enough cases to establish whether consistently at
the same long ISI it is present when the subject has no task and
absent when the subject is instructed to pay attention to the
ISIs or to count the missing stimuli.
The OSP after low frequency conditioning flashes is a large, slow, positive wave, with a peak latency usually 500-1100 ms after the due-time of the missing flash or first missing flash after the end of a train (Fig. 4). Although this range in peak latency is large among subjects (mean 1002 ±143 ms for a set of experiments with ISI 1.7-3.2 s), the latency is quite
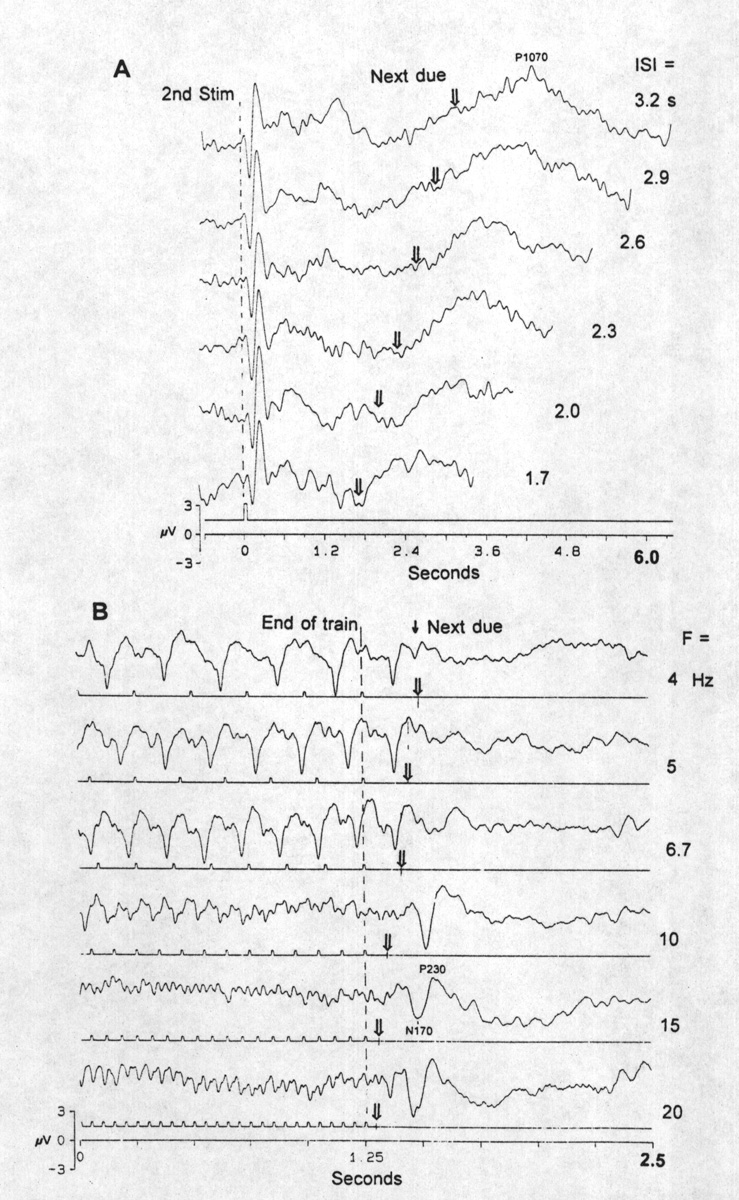 |
Fig. 4. OSPs as function of ISI, in the two ranges. A, long ISI range, 1.7-3.2 s, regular flashes, omitting every third, i.e. conditioning with a single interval. Derivation from locus P3. Each trace the average of 100 sweeps, aligned by the time of the second flash, the one before an omission. Arrows show the due-time of the missing stimulus. Compare the sharp negative-positive VEP to the second stimulus with the broad, slow, positive OSP at 800-1100 ms after the due-time. B, high frequency range, 4-20 Hz trains. Derivation from Pz; averages of 75 sweeps. Note the VEPs at 4 Hz alternate between large and small. There are only VEPs and no clear OSP <5 Hz and only the P230 (measured from the due-time of the first omitted stimulus) at 5 and 6.7 Hz; a large N170 comes in at 10-20 Hz. Bandpassed 0.1-50 Hz. |
constant for a given subject at different ISIs (standard deviation <100 ms is less than the uncertainty of measurement of these very slowly rounded maxima), as though it begins accurately at the moment a flash is expected and is then suppressed if a flash occurs. The beginning of the slow rise in positivity may have a very short latency after the due-time or even precede it, not as part of the SSR but unique to the interval during which the target is anticipated, since this is not a paradigm with random missing flashes.
We have not observed alternation in averaged VEPs (SSR) or OSPs in the low frequency range.
A3. Effects of jitter of ISI on OSP.
Since the OSP is traditionally regarded as a sign of expectation
and behaves as though the system accurately detects the absence
of a stimulus at the due-time, an obvious test is to manipulate
the expectation by making the prediction of the next stimulus
uncertain. In previously reported experiments with fish (Bullock
et al. 1990) we found that a high percentage jitter of the ISIs
around the same (6 Hz) mean as a regular train that caused a good
OSP, does not reduce the OSP substantially, as long as the last
interval is the same as the mean interval. In human subjects,
where only the low frequency range has been used heretofore, the
effect of ISI jitter has been briefly mentioned by Takasaka (1985)
who found it abolished the expectation ERP. The instruction given
to subjects in the present experiments was to try to predict the
time of the next flash; we chose to use the end-of-train method
since single omissions would be confounded by the next VEP, therefore
could not instruct subjects to count omissions.
At a mean conditioning stimulus frequency at the bottom of the high range (6 Hz), the OSP does not disappear totally with a rather high jitter, providing the train always ends with a mean ISI; this also true at the high end of the range, e.g. 20 Hz (Fig. 5A). During the irregular
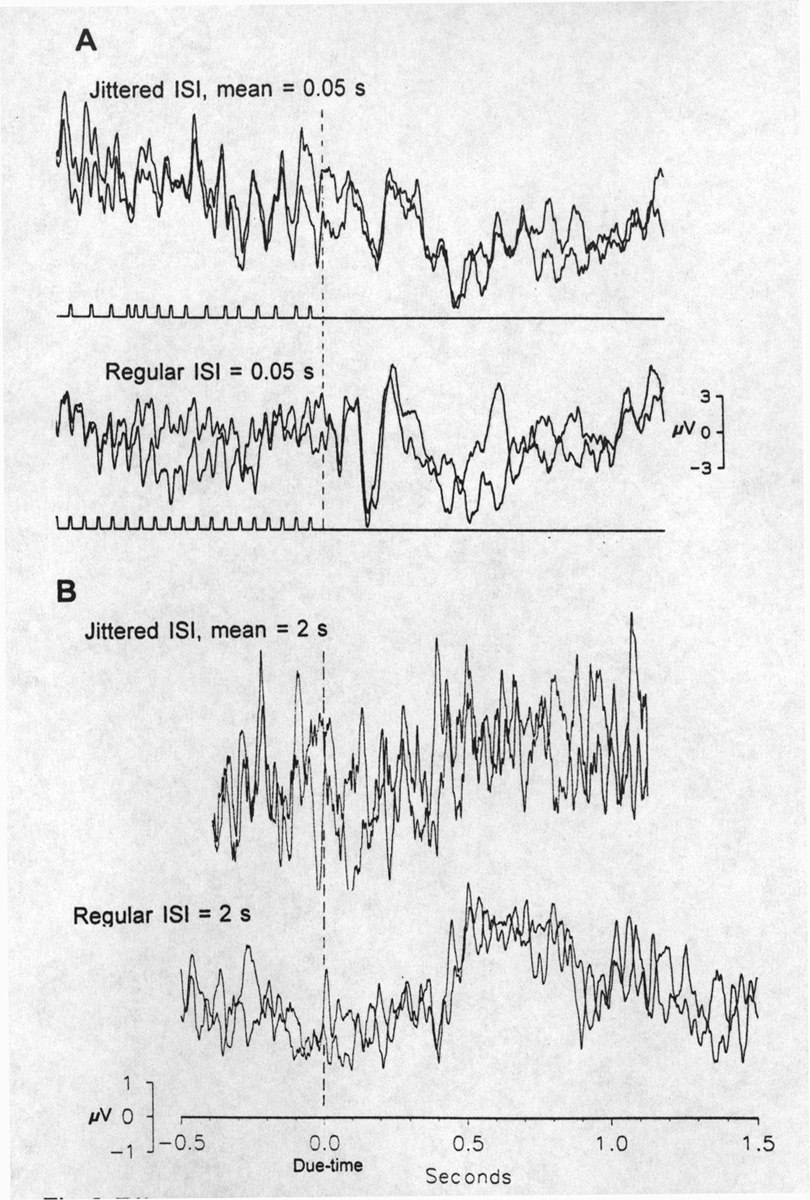 |
Fig. 5. Effect of jitter of ISIs. A, in the fast stimulation range. Below, unjittered, regular train at 20 Hz. Above, mean ISI is 50 ms; jittered between 25-75 ms pseudorandomly with the condition that the last ISI is 50 ms. Two averages of 38 sweeps, superimposed; each sweep is an identical time series of ISIs; derivation Pz. The OSP is only slightly reduced by this jitter. Note the irregular responses to the irregular stimulation during conditioning. B, effect of jitter in the low frequency range; different subject and locus. Zero time is the due-time of the omitted stimuli; no stimulus markers are shown because no stimuli occur in the period illustrated. Below, a regular series of constant ISI = 2 s, with every 4th to 7th flash omitted, causes the OSP shown, a large positive wave centered about P650. Above, when ISI is jittered around the mean = 2 s, between 1.5 and 3 s, pseudorandomly but always ending with 2 s, the OSP is not convincingly present at all. Averages of 26; amplifier filters 0.3-30 Hz; derivation P3. |
train the SSR is quite irregular, large and small VEPs following long and short intervals. We did not study in human subjects, as we have done in fish (Karamürsel and Bullock 1994), the relative importance of the last interval and of preceding intervals that can define a time constant of forgetting. This is because it would require still more averaged scalp recordings in addition to the already protracted time each subject is asked to sit.
At low mean frequencies (0.5 Hz), the OSP is wiped out or greatly reduced by jitter (Fig. 5B).
B. Effects of room light, flash intensity and stimulus type on VEP and OSP.
OSPs are not greatly affected by flash intensity, but steady state VEPs become larger with high intensities; therefore the relative amplitude of OSPs gets smaller. Especially at the low end of the high frequency range (5 & 6.7 Hz) the OSP looks like a part of a large VEP. Ambient room light slightly reduces the VEP but hardly influences the OSP. For these reasons low flash intensities in moderate room light were used in this study.
Above 3 Hz there is another illusion: The flash with its diffuser held close to the face (10-15 cm), and without near accommodation, causes an impression that the regular flashing frequency is irregular. With strong attention this illusion can be kept for a long period (ca. 1 minute). Under these conditions alternation of VEP amplitudes (every other VEP is larger) can be seen in the average (because the phase of the first alternation and the 1 in 2 ratio are consistent).
Actually, it is not important to use the diffused flash. We find quite similar OSPs with a distant flash unit (ca. 2 m), subtending only a few degrees of visual angle. It is not necessary to use the flash unit, with its 10 cm diameter and relatively bright white light. Quite similar, small OSPs and SSRs to <2 Hz can be obtained with small (3 mm) and weak light sources such as the green light emitting diode in the panel of the pulse generator that monitors the output triggers (modules of Pulsemaster A 300).
C. Effect of the type of attention on OSP
In order to show good fast OSPs (>5 Hz), it is necessary to pay attention to looking at the same point on the diffusing screen of the flash unit; fixation of eyes is important. It is not necessary to count omissions or pay attention to the frequency; in any case subjects report it is difficult to follow >3 Hz, consciously). Slow OSPs, in contrast, need some kind of conscious attention, usually assured by instructing the subject to count the omissions. OSPs to low frequencies (<1 Hz) need only attention to the frequency, not to fixation at exactly the same point. It is not even necessary to look at the flash unit; the gaze can wander so long as the subject can see enough to follow the frequency. (See paragraph above for the light requirements.) Section A2b, above described a marked effect of the instruction "Count the omissions" upon the standard VEP, reducing or abolishing the normal, large N250-350 of the VEP without attention.
D. Binocular interaction
Closing one eye or switching from one eye to the other during conditioning has no effect on the OSP at low frequencies, whereas under some conditions it can alter the SSR. Eye blink artifacts can be seen in an average without distortion of the OSP. In direct recordings from the optic tectum of rays (Karamürsel and Bullock 1994) we reported an interaction on switching from monocular stimulation of one eye to the other eye, affecting only late components of the low frequency VEP; we have not seen this in humans perhaps because it is difficult to get late components reliably from scalp recordings. We could not do interaction experiments reproducibly at high flash rates.
E. Cross-modal property of slow OSPs
Slow OSPs are independent of changing the stimulus modality during the train (cross-modal conditioning). With a specific paradigm instruction such as: "Do not pay attention to the type of stimulus, pay attention only to the frequency and try to catch the omitted ones," the same OSP can be obtained cross-modally or unimodally. At the end of the flash train, a click was added with the same ISI after the last flash, i.e. 2s. The same OSP wave sequence was obtained after the next due-time (Fig. 6), as though it were a flash, if we look at the scalp locus with the best OSP. A
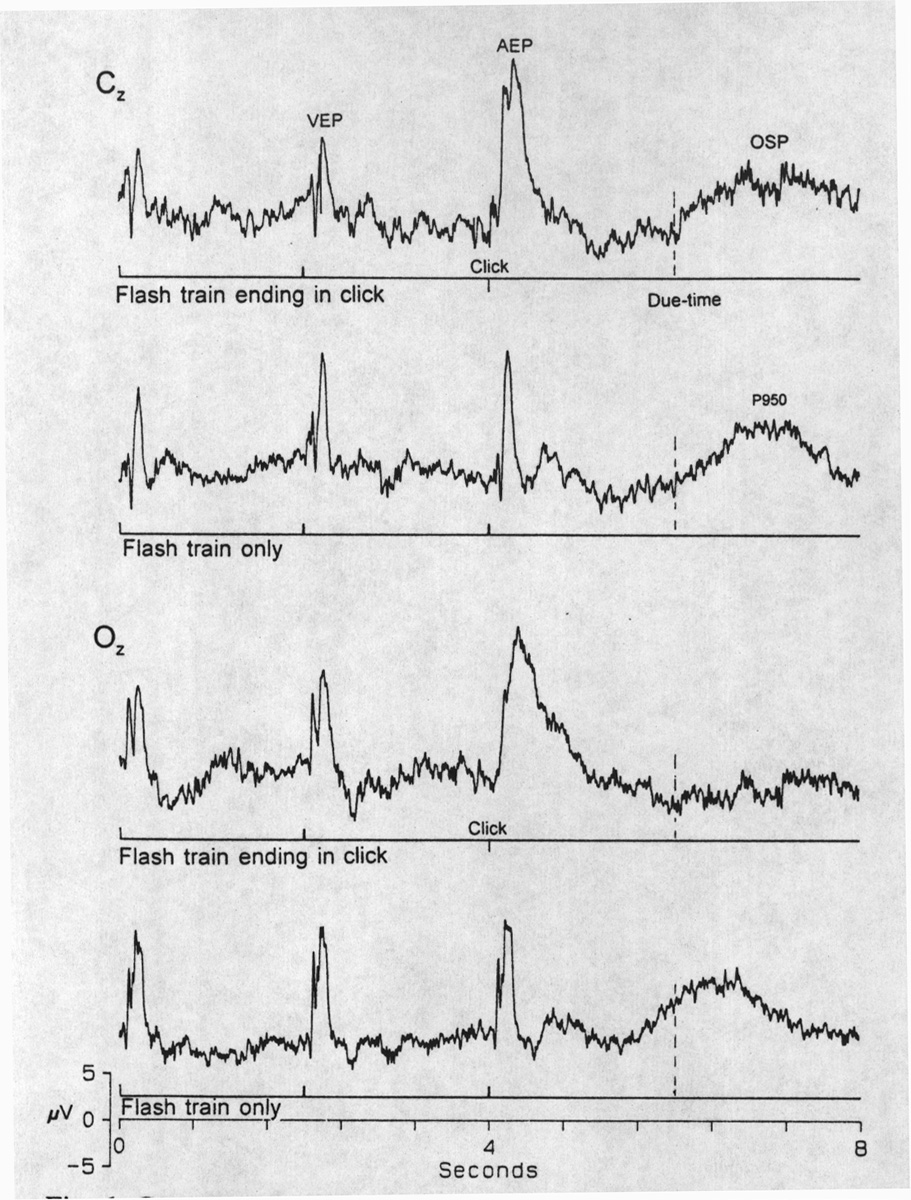 |
Fig. 6. Cross-modal OSP. The last stimulus in a regular train of 7 flashes at 0.5 Hz is changed to a click in the upper traces for each locus, Cz and Oz, but not in the lower control trace. In both loci the click elicits its own auditory evoked potential (AEP) and in locus Cz but not in Oz the click acts just like a flash in preventing the OSP. After the due-time for the next, missing stimulus, the OSP appears on schedule in Cz. Averages of 75 sweeps. |
flash after a click train generates the same result. Fast OSPs, however, are not cross-modal; a click after a flash train or a flash after click train are new stimuli that cause EPs superimposed on the OSP which comes at the time expected if the click were absent.
F. VEP and OSP in different scalp loci
The best scalp locus for the VEP is not always the best place for the OSP. The VEP and SSR are largest usually in occipital and sometime frontal derivations. The early components of the fast OSP are better in occipital while the late components are better in parietal regions and especially at the vertex (Fig. 3). Slow OSPs are generally large and clear at the vertex and at parietal loci.
G. Evidence for fluctuations in sensitivity
We do not usually see evidence for substantial fluctuations over time or recording sessions in respect to the measures studied. Similar VEP, SSR and OSP responses can be obtained from the same subject in different sessions on the same day or after several days or weeks if he/she can maintain attention. Some subjects, however, showed no clear OSP at low frequencies in early recording sessions but did in later sessions. Full attention is absolutely needed for a good slow OSP. Such attention is not necessary for the fast OSP, if fixation is maintained.
H. Induced rhythms
In four subjects, as a late part of the OSP, an oscillatory component of 10-13 Hz was observed in the averages, at 10 and 15 Hz flash rates, lasting ca. 300 ms. It should be noted that in more favorable recording conditions, for example in the turtle (Prechtl and Bullock 1994), longer lasting induced rhythms, possibly comparable to the gamma band in mammals, considering the body temperature, were seen in single sweeps but disappeared in averages due to frequency or phase jitter. It may well be that humans have such induced rhythms as a late component of the OSP, not perfectly time locked to the due-time of the omitted stimulus, and poorly seen in scalp recordings.
|
|
Discussion |
|
|
|---|
A primary finding of this research and its cognates in nonmammalian subjects is that responses in the brain to repetitive stimuli cannot be understood simply in terms of refractoriness and recovery. Processes designated - by their outcome - as facilitation, suppression, rebound and induced rhythms are some of the most obvious additional elements. They may have rates of onset and decay not only different from each other but distinct for the earlier and later components of the response and for responses to faster and slower conditioning trains.
As others have noted over the years, endogenous and exogenous events in the brain form a spectrum with intermediates. We would underline that the neurobiology of expectation - of which the omitted stimulus paradigm is a special case - covers a variety of processes, probably a variety of mechanisms, at lower and higher levels in the brain, some even quite peripheral (Bullock 1988, 1993). Since the search for an omitted stimulus ERP in lower vertebrates revealed one already in the retina (Bullock et al. 1990; Prechtl and Bullock 1994) and in medullary nuclei for a non-visual modality (Bullock et al. 1993), but only in the "high" frequency range (>2 Hz), heretofore unknown in experiments on humans, it became interesting to test for such an ERP in humans.
Another primary finding is that two kinds of OSPs are emitted after conditioning in different ranges of flash rates (Table 1). Not only do we find a missing stimulus ERP in the high frequency
| "Fast" OSP | "Slow" OSP | |
| Frequency range of conditioning flash series (large, diffuse or small, weak source) |
>2 Hz (human data: 5-40+ Hz) |
<2 Hz (human data: 0.3-1.6 Hz) |
| Latency of first main peak after due-time(4); polarity and ms | N120, P170-230, plus tendency to a late (300ms) induced rhythm (10-13 Hz) | P500-1100(5) |
| Required state | Fixation but not attention(6) | Attention but not fixation |
| Multimodal: click is equivalent to flash | No | Yes |
| Probable origin | Retina(7) | Unknown |
| Effect of jitter of ISI(8) | Slight reduction of OSP | Severe reduction of OSP |
| Required conditioning series | 250-350 ms or <5 flashes; repeated for averaging (9) |
2 flashes; repeated for averaging(10) |
| Best scalp locus | Occipital | Vertex & parietal |
| Binocular interaction | Does not exist | Does exist |
range in the present experiments, but we find this range is not continuous with or even overlapping with the conditioning frequency range used by previous authors for the familiar human ERP to missing stimuli, which is a low frequency range, <2 Hz. We have no good suggestion for an explanation of this blind spot in the spectrum but regard it as more likely apparent than real. The absence of OSPs in a range of conditioning flash rates between the "high" (>2 Hz) and the "low" (<2 Hz), actually between ca. 2 and 5 Hz is curious and invites, firstly, renewed effort to find the conditions that will reliably elicit expectation potentials - since the other two ranges show such large differences in the conditions conducive to their OSPs. Of course, absence could mean simply that there is too much latency fluctuation to survive averaging. Alain et al. (1989) used 1.7 Hz tone stimuli, at the boundary between the "low" and the absent ranges; they reported very small or unclear OSPs in nearly half their subjects; in the better 4 out of 7 subjects only some electrode loci (temporal lobe) gave OSPs, smaller on average than ERPs to targets. Their OSPs were quite unlike ours in the same range, namely P350-400 instead of the much later maximum (P500-1100) in the present report. We have not done enough with auditory OSPs to be able to say whether this difference in modality could explain the difference in latency.
In a number of dynamic properties, the two forms of OSP to two distinct frequency ranges of conditioning stimuli are quite different, including dependence on attention and on fixation, jitter of the conditioning intervals and the acceptance of stimuli of other modalities as part of the regular, expected series that prevents the OSP after each stimulus. Induced rhythms are a labile, late component of OSPs, especially of the fast type, in each species studied, sampling fish and reptiles as well as humans. These contrasts suggest that at least some parts of the mechanisms are distinct. Humans have both types; the nonmammals examined among fish and reptiles have so far revealed only the high frequency range OSPs. It is now of interest to learn whether the only obstacle to demonstrating the slow type in nonmammals is building in some attention, for example by reinforcing the omissions with classical conditioning.
Characteristic of both types is the surprisingly small number of conditioning intervals necessary to establish the temporally specific apparent expectation. Even after a single interval (two stimuli; Fig. 4A) a wave occurs at the characteristic latency for unequivocal OSPs that satisfy the defining condition: being not attributable to VEPs.
In our fish and reptiles we have found the fast OSP is already present in the retina and repeated, with modification in the midbrain, from which one interprets this ERP to be not a high level cognitive expectation. We have called it evidence of an "apparent expectation," because of the tendency to a constant latency over a wide range of ISI's (Bullock 1988, 1993), and offered an explanation based on low level properties that could even belong to receptors (Bullock et al. 1990, 1993; Prechtl and Bullock 1994). What about the slow OSP; at what level does it appear? As part of the assessment of its cognitive level, it will be important to find the means to discover whether the slow OSP begins in the brainstem or even the retina in humans. It would also be desireable to know whether the fast OSP is similar to that of the sample species of fish and reptiles in arising in the retina.
Parenthetically, it should be pointed out that we repeatedly refer to the retina only because the present study was based on visual stimuli. "Earlest afferent stations and possibly even receptors" would be more informative, since we found OSPs in another modality, electroreception, in elasmobranchs, in the first medullary nucleus and some signs of them in the primary afferent axons (Bullock et al. 1993).
VEPs are noteworthy, even without dwelling on the marked diversity among human subjects in the amplitude and form, which are already familiar in the literature, or upon the transition from the rested or long-ISI form to that of the SSR, a transition which is not yet adequately described, let alone understood. Alternation of successive VEP amplitudes (large, small, large, small ... = "1 in 2") at certain flash rates (>ca. 4 Hz), aided by conditions of attention and looking, is a striking dynamic property that we have been able to explore only in a preliminary way. We are reminded of the quite consistent alternation in elasmobranchs (Karamürsel and Bullock 1994), extending from 1 in 2 up to a large VEP every 4th and occasionally every 7th flash. Such high ratios as 1 in 7 belong in the same domain as the evoked responses to flashing between the uppermost following frequency (UFF; 1:1 without averaging) and the lowermost fusion frequency (LFF; no ripple even in the average). While numerical values for these landmarks depend on many factors, the point of interest revealed by evoked potentials is that, in this frequency zone, there is no steady state but an irregular succession of small EPs or, on occasion a regular, high ratio. We report here that alternation is not unknown in humans in the same frequency range, even in averages. A few observations are recorded on illusions seen by subjects during alternating VEPs.
Our interpretation of the fast OSP, which in our fish and reptiles begins already in the retina, is quite different from the usual view of the slow OSP. As we have proposed elsewhere (Bullock et al. 1990; Karamürsel and Bullock 1994), this "apparent expectation" might mean that the equilibrium of inhibition and excitation achieved during the conditioning train automatically makes the latency of onset of the excitatory process just barely longer than the last ISI so that if the inhibition contributed by a stimulus is missing or delayed by as little as 5-7 ms, an after-discharge process equivalent to a stimulus proceeds unhindered to consume its characteristic latent period for firing the OSP.
The SSR and the OSP represent two kinds of short term memory. We conclude that a major category of sensory response characteristics, besides the classical ones (sensitivity, receptive fields, recovery times and the like), is that of dependence upon recent history of iterative events, including their intervals, delays and omissions; the dependencies we speculate are multiple and partly independent, for example multiple facilitating and forgetting time constants. Such a conclusion means that new effort is needed to study single sweeps (Achimowicz 1991) and the relation of responses to the prestimulus or pre-event background activity. The present study has examined parametrically only some variables of recent history. Such dynamic characteristics are important neglected properties of afferent systems at each level from the receptor onwards.
Some of these dynamic properties might be found useful in characterizing the state of the brain in pathological conditions, in recovery of function after insults, and in stages of development and aging. Presumably there will be a trade off between robustness and sensitivity that will be different for example, as between fast and slow OSPs. The reason for so thinking is that these properties are probably dependent on nonlinear interactions of two or more time varying processes, sensitive to state and history as well as input. It remains to be learned whether the most useful combination of robustness and sensitivity for a particular condition will be the OSP latency or amplitude, lowest conditioning frequency, amplitude increase with conditioning frequency, induced rhythm, UFF, LFF or some derivative of these.
|
|
References/Acknowledgements |
|
|
|---|
Achimowicz J.Z. On the deterministic brain states revealed by VEP classification in phase domain. In: I. Dvorak and A.V. Holden (Eds.) Mathematical Approaches to Brain Functioning Diagnostics. Manchester, Manchester University Press, 1991: 209-230.
Alain, C., Richer, F., Achim, A. and Saint Hilaire, J.-M. Human intracerebral potentials associated with target, novel, and omitted auditory stimuli. Brain Topogr., 1989, 1:237-245.
Barlow, J.S., Morrell, L. and Morrell, F. On evoked responses in relation to temporal conditioning to paired stimuli in man. MIT Res. Lab. Electron. Quart. Prog. Rep., 1965, 78: 263-272.
Bullock, T.H. The comparative neurology of expectation: stimulus acquisition and neurobiology of anticipated and unanticipated input. In: J. Atema, R.R. Fay, A.N. Popper and W.N. Tavolga (Eds.), Sensory Biology of Aquatic Animals. Springer-Verlag, New York, 1988: 269-284.
Bullock, T.H. How Do Brains Work? Papers of a Neurophysiologist, Birkhäuser, Boston, 1993: 204-215.
Bullock, T.H., Hofmann, M.H., Nahm, F.K., New, J.G. and Prechtl, J.C. Event-related potentials in the retina and optic tectum of fish. J. Neurophysiol., 1990, 64: 903-914.
Bullock, T.H., Karamürsel, S. and Hofmann, M.H. Interval-specific event related potentials to omitted stimuli in the electrosensory pathway in elasmobranchs: an elementary form of expectation. J. Comp. Physiol. A, 1993, 172: 501-510.
Courchesne, E. From infancy to adulthood: the neurophysiological correlates of cognition. In: J.E. Desmedt (Ed.), Cognitive Components in Cerebral Event-Related Potentials and Selective Attention. S. Karger, Basel, 1979: 224-242.
Ford, J.M. and Hillyard, S.A. Event-Related Potentials (ERPs) to interruptions of a steady rhythm. Psychophysiology, 1981, 18: 322-330.
Hillyard, S.A. and Picton, T. Electrophysiology of cognition. In: F Plum (Ed.) Handbook of Physiology: The Nervous System. Vol V, Part 2, Higher Functions of the Brain. American Physiological Society, Bethesda, MD, 1987: 519-584.
Karamürsel, S. and Bullock, T.H. Dynamics of event-related potentials to trains of light and dark flashes: responses to missing and extra stimuli in rays. 1994, submitted with this manuscript.
Kerkhof G.A. Decision latency: the P3 component in auditory signal detection. Neurosci. Lett., 1978, 8: 289-294.
Klinke, R., Fruhstorfer, H. and Finkenzeller, P. Evoked responses as a function of external and stored information. Electroenceph. clin. Neurophysiol., 1968, 25: 119-122.
Mangun, G.R., Hillyard, S.A. and Luck, S.J. Electrocortical substrates of visual selective attention: synergies in experimental psychology, artificial intelligence, and cognitive neuroscience. In: D.E. Meyer and S. Kornblum (Eds.), Attention and Performance XIV. M.I.T. Press, Cambridge, MA, 1993: 219-243.
McCallum, W.C. Brain slow potential changes elicited by missing stimuli and by externally paced voluntary responses. Biol. Psychol., 1980, 11: 7-19.
Picton, T.W., Hillyard, S.A. and Galambos, R. Cortical evoked responses to omitted stimuli. In: M.N. Livanov (Ed.), Major Problems of Brain Electrophysiology. Academy of Science, USSR, 1974: 302-311.
Prechtl, J.C. and Bullock, T.H. Barbiturate sensitive components of visual ERPs in a reptile. Neuroreport, 1992, 3: 801-804.
Prechtl, J.C. and Bullock, T.H. Event related potentials to omitted visual stimuli in a reptile. 1994, submitted with this manuscript.
Renault, B. and Lesevre, N. Topographical study of the emitted potential obtained after the omission of an expected visual stimulus. In: D.A. Otto (Ed.), Multidisciplinary Perspectives in Event-Related Brain Potential Research. National Technical Information Service, Springfield, VA, 1977: 202-208.
Renault B. and Lesevre N. A trial by trial study of the visual omission response in reaction time situations. In: D. Lehmann and E. Callaway (Eds.), Human Evoked Potentials. Plenum Press, New York, 1978: 317-329.
Ritter, W., Simson, R. and Vaughan, H.G. Topographic analysis of task-related cerebral potentials. In: J.E. Desmedt (Ed.), Cognitive Components in Cerebral Event-Related Potentials and Selective Attention. S. Karger, Basel, 1979: 132-139.
Ruchkin, D.S. and Sutton, S. Latency characteristics and trial-by-trial variations of emitted cerebral potentials. In: J.E. Desmedt (Ed.), Cognitive Components in Cerebral Event-Related Potentials and Selective Attention. S. Karger, Basel, 1979a: 106-118.
Ruchkin, D.S. and Sutton, S. CNV and P300 relationships for emitted and for evoked cerebral potentials. In: J.E. Desmedt (Ed.), Cognitive Components in Cerebral Event-Related Potentials and Selective Attention. S. Karger, Basel, 1979b: 119-131.
Rusinov, V.S. General and localized alterations in the electroencephalogram during the formation of conditioned reflexes in man. Electroenceph. clin. Neurophysiol., 1960, 13: 309-319.
Squires N.K., Squires K.C. and Hillyard S.A. Two varieties of long latency positive waves evoked by unpredictable auditory stimuli in man. Electroenceph. clin. Neurophysiol., 1975, 38: 387-401.
Squires, N.K., Squires, K.C. and Hillyard, S.A. Functional equivalence of signal-present, signal-absent, and threshold-detect P3s. In: D.A. Otto (Ed.), Multidisciplinary Perspectives in Event-Related Brain Potential Research. National Technical Information Service, Springfield, VA, 1977: 218-221.
Sutton, S., Braren, M., Zubin, J. and John, E.R. Evoked-potential correlates of stimulus uncertainty. Science, 1965, 150: 1187-1188.
Sutton, S., Tueting, P., Zubin, J. and John, E.R. Information delivery and the sensory evoked potential. Science, 1967, 155: 1436-1439.
Takasaka, Y. Expectancy-related cerebral potentials associated with voluntary time estimation and omitted stimulus. Folia Psychiatr. Neurol. Jpn., 1985, 39: 167-172.
|
|
Footnotes |
|
|
|---|
1. Aided by grants from the National Institute of Neurological Diseases and Stroke, the National Science Foundation and the U.S. Air Force Office of Scientific Research.
2. 2 Supported by the Department of Physiology, Istanbul Medical Faculty, and Electro-Neuro-Physiology Research and Application Center, University of Istanbul, Turkey,
3. 3 Physiological Institute, Medical University, Lübeck, Germany. Supported by the German Science Foundation. Some of the experiments were done in the laboratory of Prof. E. Baar.
4. Latency tends to be the same for any conditioning frequency within the range.
5. There may not be a difference in the tendency to a late induced rhythm since slight jitter of its latency, phase or frequency would reduce it greatly in averages.
6. Fixation of gaze upon the light source. Attention to the timing of the omission. See instructions to subjects.
7. Based on experiments on rays and turtles (Bullock et al. 1990; Prechtl and Bullock 1994)
8. See text for details of the mean frequency and range of jitter.
9. This is not necessarily different from the slow OSP. Due to the shorter latency, five is about the minimum to satisfy the definition of an OSP - that it cannot be considered part of the last VEP.
10. The OSP appears in averages 500-1100
ms after the due-time of the missing third flash.