![]() Top
Top
![]() Abstract
Abstract
![]() Introduction
Introduction
![]() Methods
Methods
![]() Results
Results
![]() Discussion
Discussion
![]() Acknowldgement/References
Acknowldgement/References
|
|
|---|
T.H. Bullock1, J.Z. Achimowicz2, R.B. Duckrow3 and S.S. Spencer4, V.J. Iragui-Madoz1
1 Department of Neurosciences, University of California, San Diego, La Jolla, CA 92093-0201
2 Department of CNS Diagnostics, Polish Air Force Institute of Aviation Medicine, Warsaw, Poland
3 Department of Neurology, University of Connecticut Health Center, Farmington, CT 06030
4 Department of Neurology, Yale University School of Medicine, New Haven, CT 06510
Running title: Bicoherence of Intracranial EEG
Address for correspondence:
T.H. Bullock
Dept. Neurosciences 0201
University of California, San Diego
La Jolla, CA 92093-0201
Tel: 619-534-3636
Fax: 619-534-3919
E-mail: tbullock@ucsd.edu
|
|
Abstract |
|
|
|---|
The hypothesis that the intracranial EEG has local structure and short-term non-stationarity is tested with a little-studied measure of nonlinear phase coupling, the bicoherence in human subdural and deep temporal lobe probe data from 11 subjects during sleeping, waking and seizure states. This measure of cooperativity estimates the proportion of energy in every possible pair of frequency components, F1, F2 (from 1-50 Hz in this study), that satisfies the definition of quadratic phase coupling (phase of component at F3 , which is F1+F2, equals phase of F1 + phase of F2). Derived from the bispectrum, which segregates the nonGaussian energy, auto-bicoherence uses the frequency components in one channel; cross-bicoherence uses one channel for F1 and F2 and another for F3. These higher order spectra are used in physical systems for detection of episodes of nonlinearity and transients, for pattern recognition and robust classification, relatively immune to Gaussian components and low signal to noise ratios.
Bicoherence is found not to be a fixed character of the EEG but quite local and unstable, in agreement with the hypothesis. Bicoherence can be quite different in adjacent segments as brief as 1.6 s as well as adjacent intracranial electrodes as close as 6.5 mm, even when the EEG looks similar. It can rise or fall steeply within millimeters. It is virtually absent in many analysis epochs of 17s duration. Other epochs show significant bicoherence with diverse form and distribution over the bifrequency plane. Isolated peaks, periodic peaks or rounded mountain ranges are either widely scattered or confined to one or a few parts of the plane. Bicoherence is generally an invisible feature: one cannot usually recognize the responsible form of nonlinearity or any obvious correlate in the raw EEG.
During stage II/III sleep overall mean bicoherence is generally higher than in the waking state. During seizures the diverse EEG patterns average a significant elevation in bicoherence but have a wide variance. Maximum bispectrum, maximum power spectrum, maximum and mean bicoherence, skewness and asymmetry all vary independently of each other. Cross-bicoherence is often intermediate between the two auto-bicoherence spectra but commonly resembles one of the two. Of the known factors that contribute to bicoherence, transient as distinct from ongoing wave forms can be more important in our data sets. This measure of nonlinear higher moments is very sensitive to weak quadratic phase coupling,; this can come from several kinds of waveforms. New methods are needed to evaluate their respective contributions. Utility of this descriptor cannot be claimed before more carefully defined and repeatable brain states are studied.
Key words: Bispectrum; Nonlinearity; Epilepsy; Cooperativity; Subdural; Hippocampus ; Higher moment
|
|
Introduction |
|
|
|---|
The hypothesis tested is that human intracranial EEGs are generally highly differentiated and local in respect to measures of cooperative interactions, at least down to millimeter dimensions, and fluctuate in time within seconds. This view is quite in contrast to a common view, derived from scalp recording, of a more global, more widely coherent EEG with relatively stationary states often lasting for many seconds. We undertook to test the hypothesis with a measure of higher moments and nonlinear cooperativity, the bicoherence, derived from the bispectrum, introduced into oceanography by Hasselman et al. (1963).
The hypothesis was suggested by evidence from voltage/time series ("raw records") and linear spectra of EEG such as those revealed by Freeman and his associates (Freeman and Skarda, 1985; Freeman and Viana di Prisco, 1986; Freeman and Baird, 1987; Freeman and Davis, 1990; Freeman, 1992), among many other authors, and by evidence of the fine structure of coherence (Bullock, 1989; Bullock and McClune, 1989; Bullock et al., 1995a, b).
The aim of the present paper is to examine the simplest of the higher moments, the bispectrum and particularly the bicoherence, which is essentially the normalized bispectrum, since this sign of interaction varies quite independently of the linear power and coherence spectra and represents a little studied descriptor. A long-range goal is to find descriptors of dynamical properties of brain function that might discriminate between parts, states, and ontogenetic stages of the brain, and especially between evolutionary grades such as those of insects, cephalopods, fish, reptiles and mammals. The goal of seeking descriptors is modest but it reflects the conviction that our real understanding of the operations taking place in organized neural assemblies is still primitive and that natural history or "system identification" is needed. Lopes da Silva (1987) opining that higher order spectra need to be examined, states "this is particularly true for the spectrum corresponding to the second order autocovariance function ..." - the bicorrelation function whose frequency domain equivalent is the bispectrum.
The bispectrum and its normalized derivative, bicoherence, look at the components of a time series that deviate from a Gaussian amplitude distribution. Bispectra have been used to describe particular aspects of ocean waves and various physical time series (Hasselman et al., 1963; Elgar and Guza, 1985; Nikias and Raghuveer, 1987; Pezeshki et al., 1990; Mendel, 1991; Chandran and Elgar, 1993). Only a few papers have dealt briefly with the EEG (Dumermuth et al., 1971, 1983; Dumermuth and Gasser, 1978; Dumermuth and Molinari, 1987; Barnett et al., 1971; Whitton et al., 1985; Lopes da Silva, 1987; Sherman and Zoltowski, 1989; Sherman, 1993; Samar et al., 1993; Sigl and Chamoun, 1994; Kearse et al., 1994; Eckhorn, 1994), and they mainly point out the availability and uniqueness of this measure, using principally a few samples of scalp EEG. Some have reported bispectral information about relationships between harmonic and subharmonic frequency components of EEG signals (Barnett et al., 1971; Dumermuth et al., 1971, 1975; Lopes da Silva and Storm van Leeuwen, 1978). Ning and Bronzino (1989) compared the deviation from Gaussianity and the strength of phase coupling in the rat cortex and hippocampus during slow wave sleep, REM sleep and quiet wakefulness. Tang and Norcia (1996) used the bispectrum with the steady state visual evoked potential and reported much higher signal to noise ratios and more information about interactions than with classical methods.
With these limited exceptions, little information is available on the incidence, character and frequency distribution of bicoherence and the nonlinearities that contribute to it, or to indicate how stable or how local or how sensitive this measure is over time, position or brain states. A commonly negative reaction to bispectral analysis is based, we believe, on an inadequate sampling and unrealistic expectations. Possibly also it has not been explored because (i) it is hard to visualize phase coupling between three nonharmonic frequencies, (ii) the kinds of time series that produce quadratic phase coupling are diverse and (iii) are not well delimited. Such coupling, in the presence of uncoupled wideband fluctuations, is a property that can inhere, without being noticed and with different causes. Hence we considered it timely to undertake a descriptive study to test the motivating hypothesis with this tool.
We have chosen to examine a modest number of human subjects with many intracranial recordings of EEG at fairly closely spaced electrode loci (6.5 - 10 mm), both subdural and deep in the temporal lobe. The data were recorded in the course of presurgical evaluation of epileptic patients who are not responsive to medication and become candidates for surgery. They provide episodes of normal, alert EEG, sleep and seizures. Preliminary notes have been published and include some bicoherence plots during event related potentials (Achimowicz and Bullock, 1993a, b; Bullock and Achimowicz, 1994).
|
|
Methods |
|
|
|---|
The unique features of higher order spectra, whose simplest representative is the bispectrum, are used in several fields of signal processing (Nikias and Raghuveer, 1987). Our use corresponds to their characterization of the properties of nonlinear systems that generate time series. We have found the bicoherence more informative than the bispectrum itself and emphasize it in this study, together with the separate measures of skewness and asymmetry. Bicoherence measures the proportion of energy in each pair of frequency components, F1 and F2, of the specified sample of the time series, and their sum, F3, that satisfies the definition of quadratic phase coupling, i.e. the phase of F3 is the sum of the phases of F1 and F2. Since bicoherence is the proportion of the coupled to the uncoupled energy, it is normalized (by the product of the F1 power x F2 power x F3 power) and is independent of the total energy. It is reported and plotted as the bicoherence squared, a value between zero and one for each pair of frequencies, F1 and F2, over a range, in our case, of 1-50 Hz, at the resolution of the computation, for each specified epoch, for each channel (electrode). When the component frequencies, F1 and F2, come from the same time series (channel; electrode), the results are auto-spectra (auto-bispectrum and auto-bicoherence); when they come from different channels (brain sites), the results are cross-spectra (F1 and F2 from one channel; F3 from another). Equations, statistical evaluation and further details are given in references cited above, especially Elgar and Guza (1985, 1988), Nikias and Raghuveer (1987), Mendel (1991), and Sigl and Chamoun (1994).
EEGs from 16 to 80 channels were low-passed by analog filters to 100 Hz, digitized at 200 to 256 Hz, analyzed in 256-point epochs (1 to 1.28 s), using a Kaiser-Bessel window, normalized per Kim and Powers (1979), overlapped 75% by a sliding window that advances 64 points, averaged over data segments of 2-20 s with a frequency resolution of 0.8-1.5 Hz, sometimes smoothed with reduction of the resolution to 3 Hz, and plotted to a maximum of 50 Hz.
This is a further analysis of data kindly provided by the neurologists in the epilepsy programs at UCSD (V.J. I. ) and Yale University hospitals (S.S.S. & R.B.D.). Two data sets are distinguished.
(i) The Yale data came from depth probes in the temporal lobe in 4 patients. Data from three of the subjects ("C, H, W") came in 3 files from 3 seizures for each subject, recorded for ca. 30 s preictally and 30-40 s during the start and first part of the seizures. We used 8-12 depth electrodes and 5-7 subdural electrodes on the same or the opposite side. Sleep, awake and seizure periods are identified by the neurologist from the EEG. The fourth Subject ("D") is represented by six files of 3-10 minutes each, including one seizure and three episodes each of sleep and wakefulness, identified by EEG inspection. This data comes from 80 intracranial electrodes, 10 depth and 30 subdural electrodes on each side.
(ii) The UCSD data came from subdural electrode strips on the temporal, frontal and parietal lobes, from seven subjects. Each strip had a row of 8 electrodes and we normally analyzed 16 channels at a time. Most subjects are represented by one seizure and one or two episodes of sleep and of alertness, in 5-10 files of 2-3 minutes each. The data is taken from 24 hour-a-day recordings in the ward, not under standard EEG laboratory conditions. Much of the interictal sleeping and waking EEG has isolated spikes.
Plots and evaluations are based on 11 subjects and 55 files. (The term file here means a recording of a continuous 1-10 minute sample in some identified brain state or transition, employing 16-80 electrodes.) The raw EEG records, as voltage vs time plots, were characterized separately and marked by the neurologists to identify the brain state and the start of the electrical seizures. The 3D plots show auto- and cross-bispectra, bicoherence and amplitude spectra for two channels, usually adjacent loci. For auto-spectra, the bifrequency plane is redundant and symmetrical around the diagonal of equal frequency except as values are rounded differently by the x and y display routines. Cross-spectra have been computed for pairs of adjacent electrodes only. Cross-spectra are not redundant and can be asymmetrical around the diagonal since F1 and F2 come from one record and F3 from a different time series.
A bicoherence plot on the bifrequency plane can be evaluated and compared with other plots in different ways, besides visual inspection. Two of the simplest are the maximum value of the highest peak and the mean of all values for all intersections (ca. 2500, at 50 x 50 and 1 Hz resolution). The first underrates the common case of widespread, low mountains; the second underrates the case of one or a few narrow peaks. We have not undertaken to divide the plane into areas, like compound frequency bands in order to average over the high-high pairs and the high-low pairs and the theta-alpha and other pairs, or to present slices of the bifrequency plane along various transects. These would no doubt be insightful. With a full page of six 3-dimensional plots for each 4 or 9 or 17 s segment, each pair of electrodes, and each file, for the 11 subjects, there are >>500 pages of this one kind of readout, before considering the statistical assessment of means and other analyses.
Statistical evaluation begins with estimation of the bicoherence to be expected from a stochastic data sample of the same length and power spectrum. Following the procedures of Elgar and Guza (1988) and Elgar and Sebert (1989), a subprogram converts all values not significantly above the chance level to that value; hence they are plotted as a smooth plateau representing the 95% significance level. All peaks that rise above it are considered to be significantly different from the null hypothesis of no bicoherence. The EEG epochs judged to have no appreciable bicoherence, i.e. only a few low, scattered ant-hills rising as single points above the plateau level of 95% significance (0.11 in our usual conditions) have a maximum single value rarely above 0.15 and a mean bicoherence of all intersections of ca. 0.04 (calculated without the conversion to a plateau level); these are the numbers attributable to chance with the particular departures from Gaussianity in our time series. A mean bicoherence of 0.045 or more is usually associated with obvious broad mountains or multiple narrow peaks of highly significant but only moderate bicoherence and is likely to show a maximum bicoherence of 0.25 or more, depending on the distribution of significant intersections.
We also measured some other variables (Table I): the maximum bispectrum peak, maximum peak and total area in the amplitude spectrum, total skewness (non-equivalence of EEG waves around the horizontal, time axis, averaged across epochs and electrodes, calculated as the sum of the real components of the normalized bispectrum) and total asymmetry (of EEG deflections around the vertical or voltage axis, calculated as the sum of the complex components of the normalized bispectrum); the last two are taken either as absolute magnitude, discarding the sign or without discarding the sign.
Data reduction includes the following. (1) Bar graphs condense the many 3D sheets, each for two loci, into a single 3-D graph, with a time axis, for each of the files (usually 60-120 s of EEG from 17 seizures, 15 awake episodes, and 23 sleep episodes). Each bar shows the mean of one of the measured variables for one electrode and 17 s epoch. To avoid congestion we group 8-12 electrodes and a succession of 6-8 epochs (90-120 s) into one figure for that file, subject, brain state and part of the brain. A montage of six such variables on a sheet can represent, for example, one seizure and its preictal epochs, permitting comparison with other places, times or brain states. Although this has reduced the data to manageable numbers of plots, it blurs the distribution of the bicoherence in frequency space; therefore we usually add the following step.
(2) Contour or topographic 2-D plots of the bifrequency plane with pseudocolor or shading showing the elevations of bicoherence, each represent an electrode locus and a time segment (ca. 2-20 s). They reveal the distribution of peaks in the bifrequency plane (F1 from 0-50 Hz by F2 from 0-50 Hz, with 1 to 3 Hz resolution). We arrange a number of plots of several loci (in columns) and several successive segments (in rows) on one sheet as an array of 2D plots mapping space and time, e.g. for 6 loci 10 mm apart and 5 epochs of 9s each.
(3) Statistics are computed, for each of 8 variables, including 95% confidence intervals for fluctuations around different mean bicoherence values (Elgar and Sebert, 1989 and S. Elgar, personal communication), for each of the files (brain states, episodes, subjects). Correlations between the variables are computed, to see which are closely covarying and which can vary more independently.
A number of experiments were conducted with artificial data of known composition to check our programs, test for leakage of energy into adjacent frequencies and learn the sensitivity of the displays to relevant parameters and proportions of noise. In the extreme case of just one intersection having quadratically coupled waves and a pink noise level 5 times larger (signal-to-noise ratio = 0.2), the bicoherence at this intersection was 0.15; the 95% significance level was 0.046 under those conditions. Skewness was 5 times and asymmetry was 2.6 times larger than the 95% significance level. Thus the method is quite sensitive to phase coupled signals much smaller than the "noise", i.e. the activity which is not quadratically phase coupled. This can explain why similar looking EEG records may differ in that one shows bicoherence and the other lacks any appreciable bicoherence. In another test, one frequency (e.g. 13 Hz) was amplitude modulated by another (e.g. 3 Hz) with pink noise at signal-to-noise ratio = 50. No bicoherence results unless some of the modulating frequency is added to the AM (amplitude modulated) signal; in this condition 1% AM gives a high bicoherence of 0.8 .
|
|
Results |
|
|
|---|
Our observations are arranged in the following way. Bicoherence is the principal focus. Bispectra, which combine into one mixture the bicoherence, skewness, asymmetry and absolute power, are relatively neglected in this report. We first describe the auto-bicoherence of normal, alert subjects, its temporal stability and spatial differentiation. Then we report findings on the cross-bicoherence spectra. Finally, the three brain states and major brain regions are compared.
Auto-bicoherence spectra of normal, awake EEG
In some _2 min samples of EEG, from many to most of the spectra computed for successive 9 or 17 s segments show virtually no bicoherence. This is important, not only as a finding: quadratic phase coupling is not typically or generally present. It is also a good control against prevalent, spurious sources of bicoherence. In every such sample, however, a number of segments have significant elevations somewhere in the bifrequency plane. For example in Subject K, file 2 (right temporal cortex; awake), 85 out of 128 spectra from the eight 17 s segments in 16 electrodes show little to almost no bicoherence, while 43 spectra have substantial elevations (Figs. 1, 2).
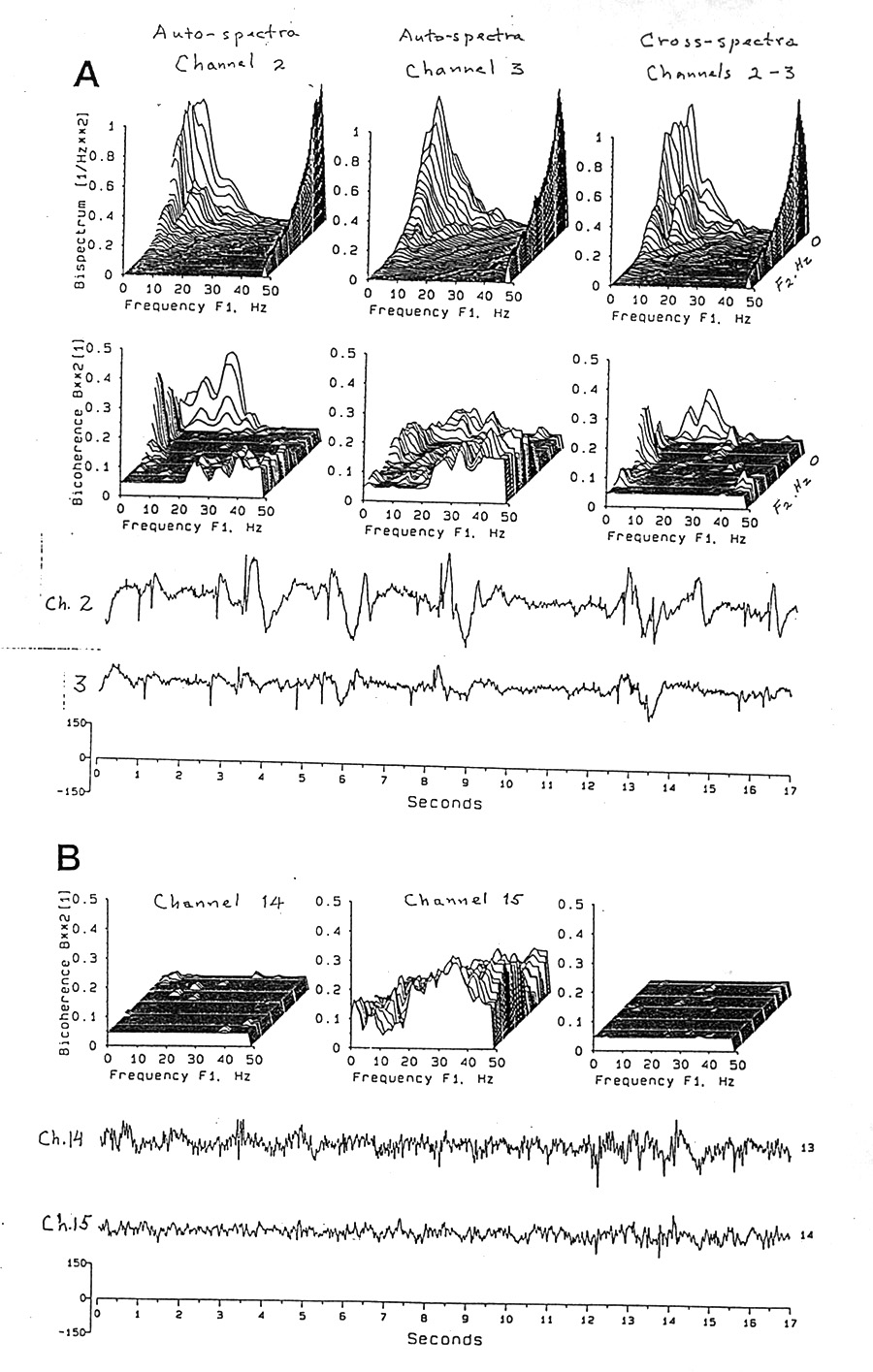
|
Figure 1. Bispectra and bicoherence spectra of EEG recorded from subdural electrodes, plotted on the plane of F1 and F2. First and second columns are auto-spectra of two electrode channels; third column shows cross-spectra between them. A, Upper row of 3-D graphs, bispectra and, as profiles on the right edge, ordinary amplitude spectra. (The units of the bispectrum are arbitrary; the dimension is volts3/Hz2 each spectrum is plotted to its own maximum, as also true for the amplitude spectra.) Lower row of graphs, spectra of bicoherence, squared. The dimension is the proportion that quadratically phase coupled energy at each intersection plus the energy at F3 whose phase is the sum of the phases of F1 and F2 bears to the total energy at these three frequencies. Values are estimated by computing for each intersection at the resolution of 1.5 Hz, from 256 points at 200 points per second (= 1.28 seconds of EEG) and repeating 50 times, advancing 64 points each time (= 75% overlap, before averaging; total 3392 points = 16.96 s). Bicoherence values of probability p >0.05 are converted to the 95% significance level to make a plateau. Note that these loci and this 17 s segment show moderately strong bicoherence in certain areas of the bifrequency plane and that these two adjacent loci have distinctly different distributions of auto-bicoherence peaks and mountain ranges on that plane. Cross-bicoherence is more like channel 2 than 3. Such plots were made for each pair of adjacent electrodes (1 & 2, 2 & 3, 3 & 4, ...) and each segment of time for the EEG samples of 1 to 6 min. Subject K, awake, right temporal cortex, subdural. Raw EEG at the bottom. B, the same Subject, asleep, right sub-frontal cortex, bicoherence only, electrodes 14 and 15. Note that these two electrodes have, respectively, virtually none and quite strong auto-bicoherence. This difference can be due to less quadratic phase coupling in locus 14 or to more wideband Gaussian fluctuation diluting it. Casual inspection cannot decide. Cross-bicoherence again resembles the lefthand autospectrum, perhaps because it contributes both F1 and F2 while locus 15 contributes only F3. Raw EEG at the bottom. |
In subdural recordings auto-bicoherence plots can be virtually flat, meaning no bicoherence, or have low, scattered ant-hills or one or a few prominent peaks or great mountain ranges. Elevations can be widespread or confined to the high-high corner (high F1 and high F2) or to the low-low corner or to the mid-F1 and mid-F2 region or the mid-high or mid-low or any region. Often there is only one main mountain but there can be two with a deep valley; sharper peaks can be single or several or many (following paragraphs). Variety of pattern is the main finding. In one example (Subject K, file 2) only one or two plots have a single peak of >0.2 height (these are duplicated in our displays by the diagonal symmetry). Overall auto-bicoherence (total area under the 3D curves) in a number of sites is high (15 out of 72), some are very high (7/72); 6/72 are low; 1/72 is very low. The rest (43/72) are distributed among the medium grades, of dubious or lower significance, statistically.
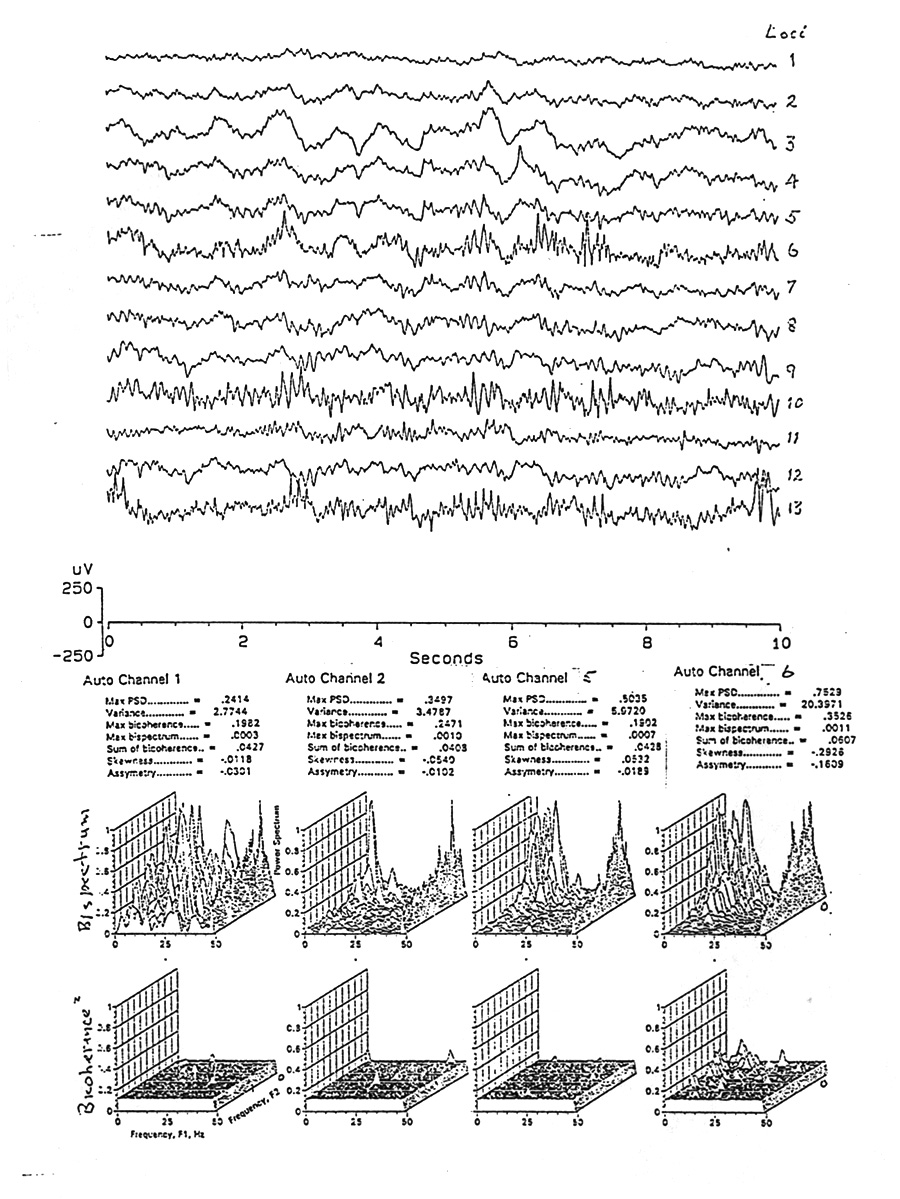 |
Figure 2. Raw EEG from 10 s of sleep in Subject H, in 13 depth electrodes (EEG) in a row horizontally through the temporal pole, amygdala and hippocampus, plus the auto-bispectra (with amplitude spectra on the right edge) and the auto-bicoherence spectra for electrode channels 1, 2, 5 and 6. Note that there is virtually no bicoherence in three of these channels, and only a few small peaks in channel 6. Bispectra are more difficult to read because they include absolute power and all forms of non-Gaussianity and they vary by orders of magnitude. Values for 7 of the measurements recorded are shown in fine print ("Max PSD" = highest peak of the amplitude spectrum; "Variance" = total area of the amplitude spectrum; Sum of bicoherence" is the mean of all bicoherence values for all intersections before converting to the 95% significance plateau; "Skewness" and "Asymmetry" are the algebraic means for all frequencies.) |
In deep temporal lobe recordings, including the hippocampus, bicoherence values can exceed 0.8 of the maximum possible in the highest peaks; mean values over all the bifrequency intersections are usually <0.1 but can be highly significant even when <0.06, as explained above. Illustrating a different method of counting than the previous paragraph, in an example from Subject C (preictal 30 s just before seizure 2), of the 12 electrodes, numbers 3, 8, 11 and 12 had virtually no auto-bicoherence (means = 0.044 to 0.052); no.9 was very small (0.062); nos.1, 2 and 4 were considerable (0.082, 0.071, 0.085, resp.); nos.5, 6 and 7 were relatively high (0.13, 0.094, 0.068 with a maximum peak of 0.59); and no. 10 had the highest mean (0.13). This subject shows power spectral peaks at 7, 14, 21, 28 and 35 Hz and bicoherence peaks in some channels but not in others.
Among the variety of patterns, 3 or 4 of 72 auto-bicoherence plots in this file of this subject show the "bed of nails" or hairbrush pattern (Figs. 3, 4). Narrow peaks have a constant spacing of ca. 6-10 Hz, filling most of the bifrequency plane but sparing the high-high corner - 1 high peaks on each side of the diagonal axis of symmetry. They may or may not correspond to the 3-7Hz spikes in the raw EEG. More striking instances of this periodic pattern were found in some other subject's files, e.g. "H," correlating with regular seizure spikes at the same frequency in the raw data. Another occasional pattern is a ridge of elevated bicoherence parallel to the F1 axis, meaning quadratic phase coupling between a certain low frequency and all other frequencies at least to 50 Hz.
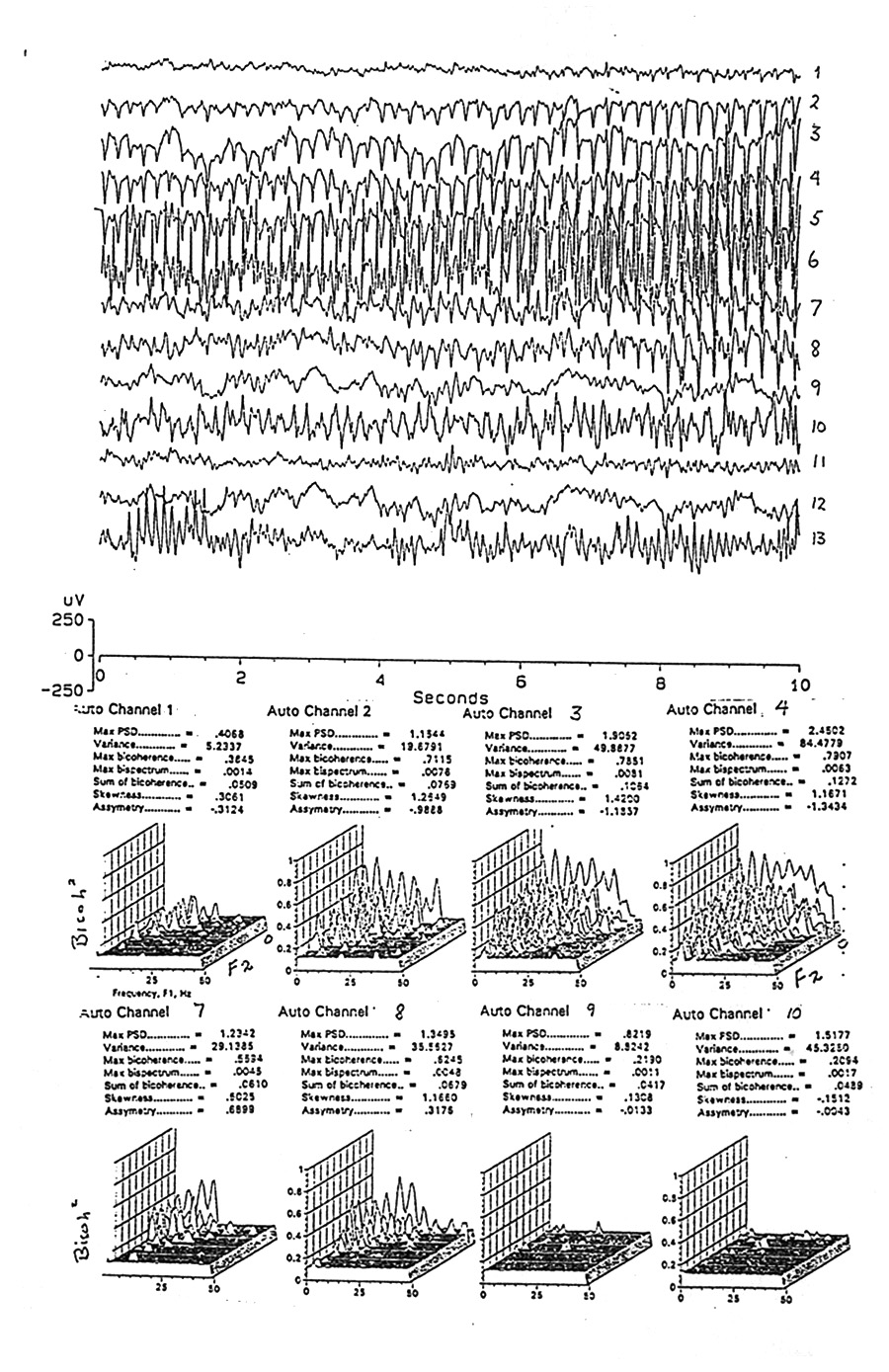 |
Figure 3. Raw data from an early part of seizure 1 in Subject H (ca. 0.5 min after Fig. 2), plus the auto- bicoherence spectra for the same segment for 8 of the channels. Channels 9-12 have virtually no bicoherence although channel 10 might be expected to. The highest spectra have a "bed of nails" pattern with a 6 Hz modulus, covering a wide area of the bifrequency plane,.but sparing the high-high corner This is not the same as the conspicuous 4 Hz waves in the EEG. This periodic pattern of bicoherence peaks has been seen only in a few seizures. |
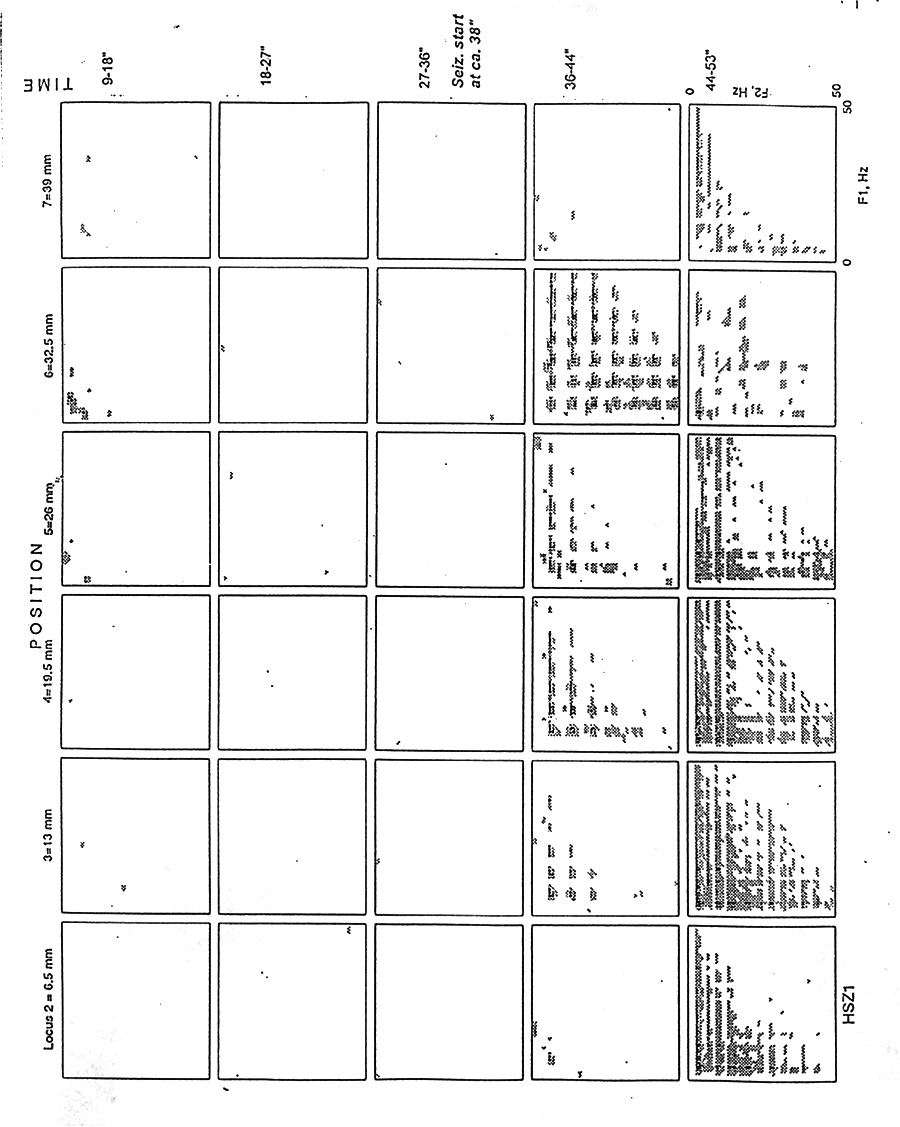 |
Figure 4. Topographic plots - like the 3-D plots of previous figures, viewed from above - of six adjacent electrode loci and five successive epochs of 9 s each, to show the distribution of auto-bicoherence in the bifrequency plane and along the length of the depth probe at 6 mm intervals and in time. Subject H, seizure 1, same data as Fig. 2; note the symmetry around the diagonal from upper left (low-low frequency) to lower right (high-high frequency). White = values below the 95% significance level; darkest gray = 0.6. Note only occasional small hills in the low-low corner in a few loci before the seizure starts in the fourth row and brings extensive bicoherence with the "bed of nails" pattern. |
Table I shows the means and 95% confidence intervals for a variety of measures in the waking state, pooling a number of electrodes and time segments. Visual inspection suggests apparent correlates of strong bicoherence in the wave forms of the raw EEG (voltage vs time = V/t plots) in some plots but other segments show that these apparent correlates are not real. A crude way of testing this is to compare two adjacent channels (electrode loci), with respect to the overall mean of bicoherence, e.g. approximately equal or greater in one than in the other, and then independently estimating by visual inspection of the activity in the raw EEG V/t plot, some candidate or suspected feature. A representative example is K02 (awake, with interictal spikes in all channels): the first segment of 17 s, has 5 strikingly opposite trends of the two estimates out of 14 pairs ("5/14"), i.e. one increases and the other decreases; segment 2 (seconds 17-34) has four; segment 3 has 5; segment 4 has four - in sum, 18 out of 56 change in unexpected directions. The majority that change in the same direction may mean there is a weak tendency for positive correlations of raw EEG features and bicoherence, but more importantly, the strong minority indicate that visible features are not reliable indicators of bicoherence. Instead of comparing simultaneous channels, we also looked at changes in the same electrode in separate time segments. In 15 out of 45 the pairs are conspicuously divergent in direction of change. It appears to be impossible to find a consistent relation between time domain EEG pattern, by inspection, and bicoherence pattern.
|
|
Table I. Sample of auto- and cross-spectra for selected measures in Subject "D" in two waking episodes of 3 min. each. Each value is the mean (large numbers) or 95% confidence interval (small numbers) for 20 epochs of 9 s each, pooled from 10 electrodes 6.5 to 10 mm apart in one of the 8 brain regions: rpt, right posterior temporal depth probe, traversing amygdala and hippocampus; lpt, same on left; rfp, right frontopolar subdural strip; lfp, same on left; rat, right anterior temporal subdural strip; lat, same on left; rpo, right parieto-occipital subdural strip; lpo, same on left. mxp, maximum of normal amplitude spectrum, arbitrary units; mxbisp, maximum of bispectrum (1/Hz2); mxbico, maximum bicoherence, squared; mnbico, mean bicoherence, squared (value for no-significant bicoherence is ca.0.03); skew, skewness, incl. signs; asym, asymmetry, incl. signs. Note: the two depth probes and eight subdural strips have significantly different mean bicoherence and some other measures, with some significant differences between episodes "Awake 1" and "2". Cross-spectra show almost as much bicoherence as auto-spectra but much more skewness and asymmetry. N = 207 for each value. |
Neighboring channels that show large differences in bicoherence without any discernible basis in the EEG are illustrated in Figure 5. This is especially true when the bicoherence mountain is in the high-high corner (>30 Hz), because we cannot see this component in the EEG, since it has such low power.
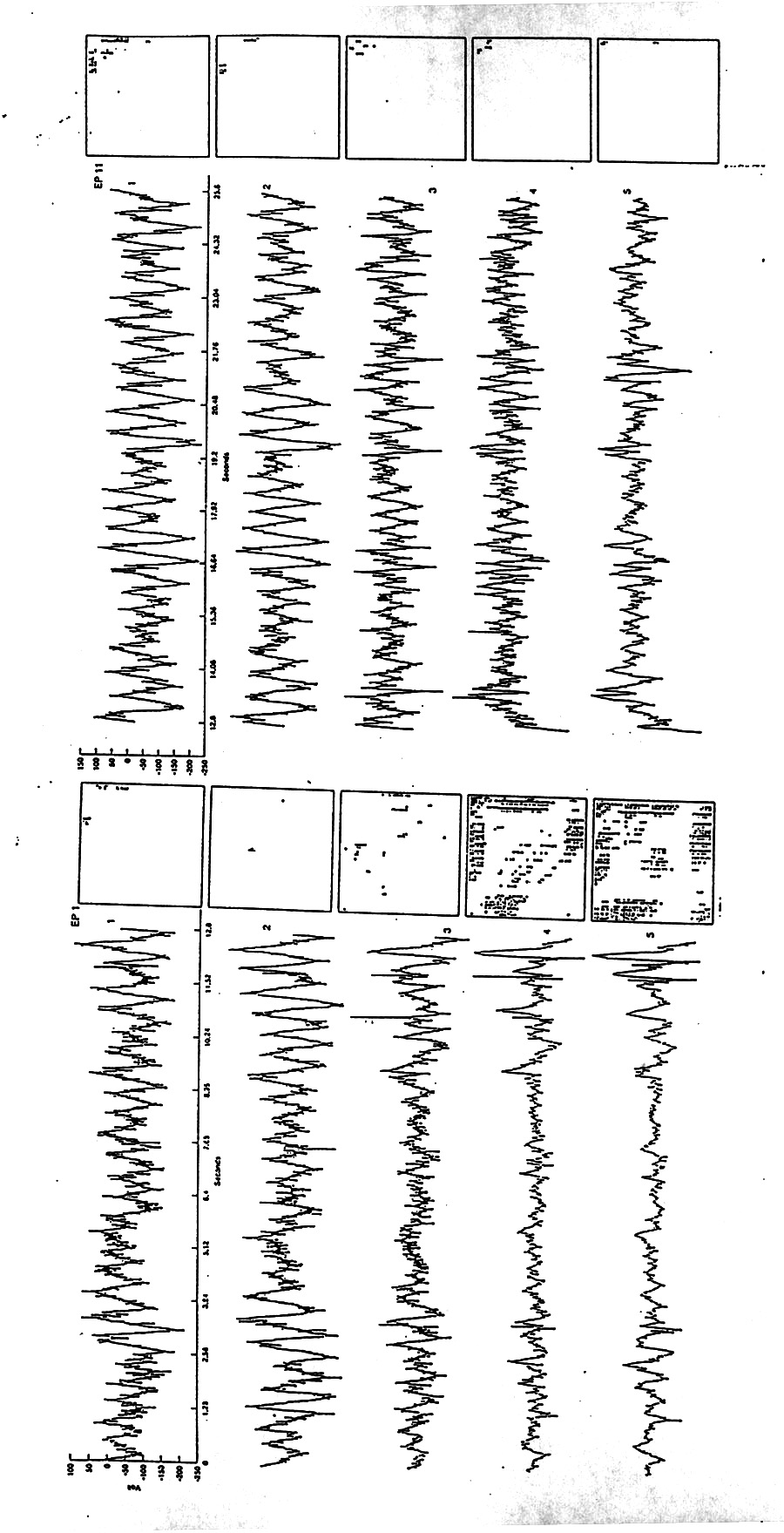 |
Figure 5. Raw data and topographic plots for a 13 s segment for the first 5 electrode loci in the right deep temporal lobe probe in Subject D (10 mm apart, #1 in the tip of the temporal pole in fusiform grey matter,#2 in fusiform white matter, #3 in amygdala, #4 and #5 on the ventricular surface of hippocampus). Awake, session 1, with a few interictal spikes and large delta waves in the temporal pole. See Fig. 4 for explanation of plots; they are rotated here so that the low-low frequency corner is lower, left. Note that adjacent electrodes can have almost no auto-bicoherence in one and extensive mountain ranges of auto-bicoherence in the other (loci 3 and 4) - with no obvious difference in the V/t record. Similarly for successive epochs, the next 13 s epoch in channel 4 (not shown) appears spikier and perhaps more asymmetrical but has only the tiniest hill of bicoherence in the low-low corner. Once more: such differences can mean changes in the absolute amount of quadratic phase coupling or in the uncoupled fraction; in the latter case this would have to involve all frequencies, which is unlikely in Fig.5. |
We have looked at the measures of asymmetry and skewness in the raw EEG, as well as spikes and other abrupt transients. Sometimes, as in the intense electrical seizure of files Csz2 and Csz3, a lot of conspicuously asymmetric and skewed waves do not give more than moderate peaks of bicoherence. This of course, can mean that an even greater increase took place in the uncoupled components, providing this involved the same frequencies, i.e. was quite wideband. In view of the signal to noise ratios tested with simulated data, and the statements of Elgar and Sebert (1989) on the relative immunity of bicoherence to background noise, this explanation is unlikely. In some channels it appears as though one or two large spikes, skewed (around the voltage axis) can make a large bicoherence over many intersections of the bifrequency plane, but we have not tested this hypothesis, for example by surgical excision of these spikes from the time series.
The table of correlations (Table II) compares each variable with others in nine data sets. Note that four classes of pairwise correlation are clear. (i) Some show a consistently high correlation, e.g. the height of the maximum amplitude peak and the total amplitude variance in all frequencies. (ii) Some show a medium to high correlation, varying by several tenths in different data sets, e.g. mean bicoherence and maximum bicoherence, mean bicoherence and asymmetry, mean bicoherence and skewness. (iii) Some show only a low to medium correlation, e.g. maximum bicoherence and maximum bispectrum, maximum amplitude and maximum bispectrum; and (iv) some are extremely variable, fluctuating from very high to very low, e.g. mean bicoherence and maximum bispectrum, mean bicoherence and total amplitude variance, skewness and asymmetry. The principal finding is that, although correlated on the average, variables such as bispectrum, bicoherence, skewness, asymmetry and power are essentially independent and do not predict each other.
|
|
Table II. Correlations of some pairs of variables in a sampling of data sets. A selection of 25 of the 182 possible pairs of measured variables, arranged in the order of their mean correlations. Data sets from four subjects: H, W and C with deep temporal probes, include 30 s of preictal and 60 s of ictal recording for three seizures in each. Some are single files (file names with numbers), representing single seizures, to illustrate consistent correlations of some pairs of variables and inconsistent correlations for others. Columns without numbered names are pooled from three episodes in each subject. Subject K is an example of the patients with subdural electrodes during a waking period (K02) and a seizure (K04). Most sets include 9-15 unselected electrodes; the two columns with "sel" in the file name include only those electrodes that show seizure activity; this does not notably affect the correlations. Skewness and asymmetry can be either positive or negative; the sign was retained in computing means for Subject K but discarded for H, W and C. Since most values are positive, this only raises the mean fractionally and better represents the departure from Gaussianity although it destroys some of the variability, assures the correlations are positive and seemingly more significant. |
Time course of bicoherence
The part of the motivating hypothesis concerning fine temporal structure was tested by computing the bispectrum and bicoherence for epochs of various lengths from 1.6 to 17 seconds, then plotting the successive values vs time. Short segments raise the threshold of significance or "bias" plateau proportionally to the segment length (under our usual conditions 0.11 with 10 s, 0.22 with 5s, 0.34 with 3.8 s). However, the maxima of peaks also rise almost as much. Time segments of 1.6 s are of limited use for evaluating changes with time, given our usual computing epoch of 1.28 s, since in addition to the rise in the significance plateau, the rise in span of the 95% confidence interval for fluctuations around the presumed true bicoherence, or its estimate, the mean value, makes no value significantly different from the mean or from any other.
If bicoherence actually changes more slowly than the sample duration (segment length), each maximum and minimum would be defined by more than one point. In fact we find even at the shortest useable sampling time (3.8 s) that peaks and valleys generally consist of a single point and the highest peaks and deepest valleys are significant. This means bicoherence can fluctuate widely within a few seconds. EEG segments that look stationary to the eye in the V/t record can be highly differentiated in respect to quadratic phase coupling. Peaks are higher and more numerous and the average over many points is higher for the 3.8 than for the 7.6 and 15.2 s segments .
Thus it is common to see low and high bicoherence in adjacent time segments of the same channel, of whatever length, even the shortest. Apparent rhythmicity at ca. 5-10 s periods is common when the segment length is long (9-17 s), but usually disappears with shorter samples.
Separate samples of the same state (tens of minutes or hours apart) can be similar in the range of bicoherence statistics and patterns or they can be distinctly different. This is treated further, below, under Brain states.
 |
Figure 6. Bicoherence fluctuation with time. Maximum bicoherence anywhere in the bifrequency plane for a certain electrode analyzed in time segments of three lengths, from a two minute record during a seizure; Subject D. Ordinates are not to the same scale. Note the two 95% statistical levels: the 95% significance level gives a bias level that would contain 95% of the chance bicoherence values in stochastic time series. It tests the null hypothesis that there is no real bicoherence. For the three curves here, these levels are, from top to bottom: 0.33, 0.14, 0.06. The other, shown as "error" bars, gives a 95% confidence interval within which fluctuations around the true value, or the mean as an approximation, are expected by chance. It tests the hypothesis that there are no real departures from the mean. The "error" bars are for the one mean value and for 18, 42 and 100 degrees of freedom, from top to bottom, interpolated from values given in Elgar and Sebert (1989). Most fluctuations are insignificant but some suggest that beyond the chance fluctuations, bicoherence can change abruptly - within a few seconds. This is not expected if the main cause of quadratic phase coupling is an ongoing or slowly changing wave form, such as a skewed or asymmetrical wave. A wide variety of curves is found, some with apparent oscillations, some with a single cycle of a slow wave with a period >1 min. |
Spatial differentiation
Bicoherence can be very local. Although the majority of neighboring electrode sites are quite similar at a given moment, low and high bicoherence plots are often side by side, in adjacent channels (Fig. 5). For example, in "K" 7 out of 32 examined in this way are different in mean bicoherence by 20% or more. Interstrip pairs are no more likely to show such abrupt differences (8/32) than pairs within the same strip.
Influence of distance
To examine the relation between bicoherence at a given locus and that at other loci closer and farther apart, we chose to compute the correlations between each pair of loci in rows of ten electrodes in each subdural strip or depth probe. The value chosen for each locus was the mean bicoherence value over the whole bifrequency plane for a number of successive data segments in a record with 4.9 min of EEG. The data segments could be 15.36 s long and hence 19 numbers in sequence, computed for correlation as they fluctuated up and down. Or they could be 3.84 s long and hence 76 numbers in sequence; or they could be intermediate in length. This method overlooks differences in detailed pattern of bicoherences in successive time segments. Another method would have been to compare the means of the cross- bicoherence spectra for pairs of different separation. This could prove to give quite different results from the correlation method. Fig. 7 shows correlation plots of several deep temporal and subdural strips during sleep.
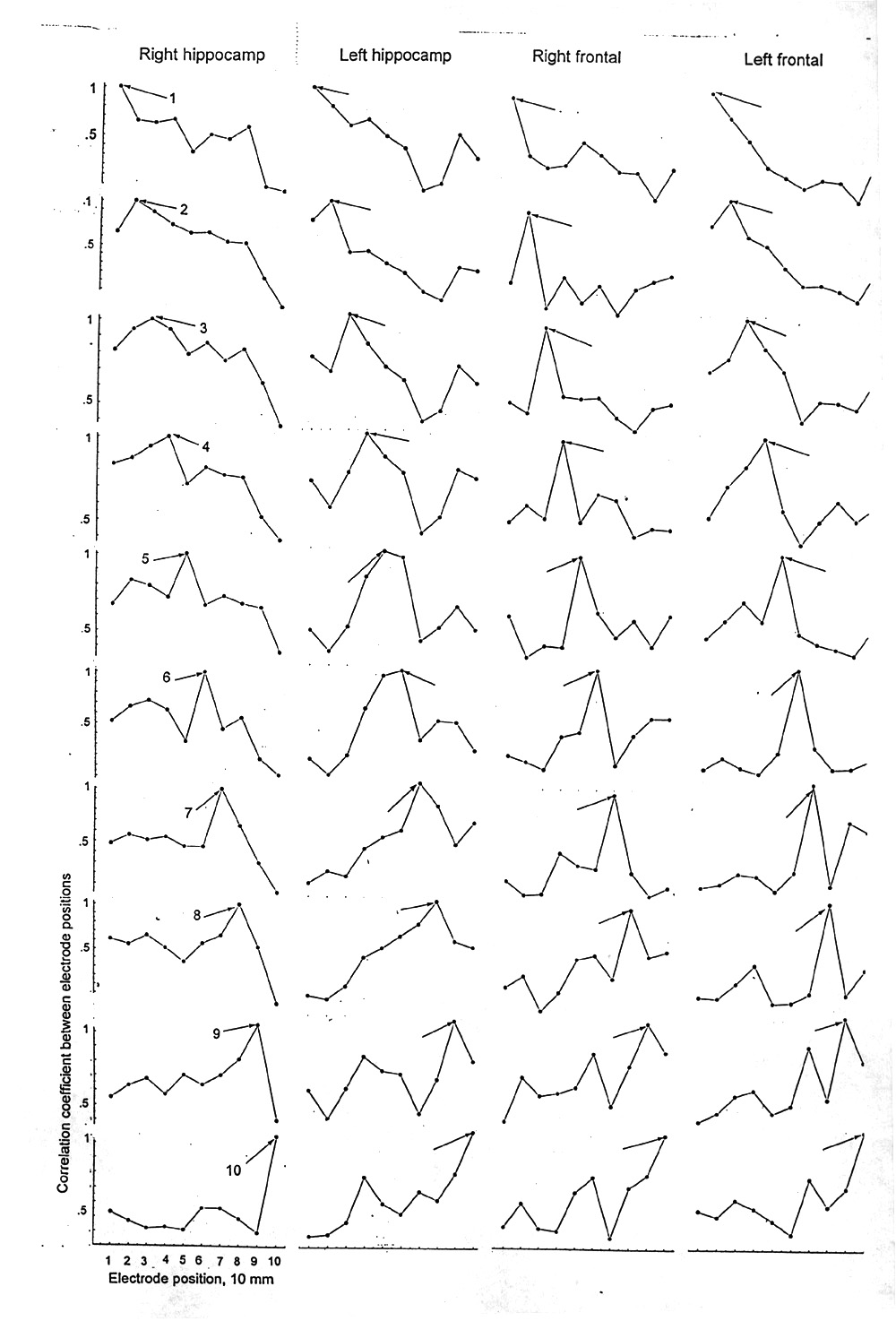 |
Figure 7. Correlation between loci vs interelectrode distance. Columns each represent a temporal lobe depth probe or a subdural strip of 10 electrodes, 10 mm apart, in right and left, approximately symmetrical sets. The abscissa is electrode number = position in the row. Each curve plots the correlations between one of the electrodes and each of the others, from closest to farthest. The common electrode is indicated by an arrow (numbered in column one) and, being self-correlated, is always 1.0. The ordinates are not to the same scale. The value for each electrode locus was the correlation coefficient of 19 successive means of the bicoherences for all intersections over the bifrequency plane for each 12.8 s data segment in a record with 4.9 min of sleeping EEG; Subject D. Note that although the average behavior is a certain slope falling with distance, a large variability is manifest between the individual pairs and often the highest correlation, apart from the self-correlated point, is not the closest pair. These curves, too, fluctuate with time. |
Typically there is a decline with distance, falling to half in about 10-30 mm. This estimate is made ignoring the 1.0 peak of self-correlation. Some curves are even steeper and some cannot be reliably distinguished from a plateau. Deep valleys of low correlation can be 20-30 mm wide. Peaks of similar width can occur at any distance within our range of 90 mm. Exceptionally we see a secondary maximum at long distances, after a steep valley. When we calculated for more numerous short time segments (3.84 s), the curves were not more jagged but quite similar to the 15 s curves.
Cross-bicoherence spectra
Cross-bicoherence has only been examined between adjacent electrodes and only for the difference interactions, where F1 and F2 come from one channel and F3 from the other. Such spectra are found in each of three categories. (i) They can look roughly intermediate between the two auto-bicoherence plots, in the distribution of peaks over the bifrequency plane. (ii) They can be lower in overall bicoherence than either of the auto-bicoherence plots. (iii) Quite commonly the cross-bicoherence closely resembles one or the other auto-bicoherence channel (Fig. 1), usually the one providing F1 and F2 and the specific pattern of the plot differs, depending on which file of the pair provides F1 and F2 and which provides F3. Cross-bicoherence is never higher than either of the auto-spectra.
Frequency-specific examination of cross-bicoherence shows great variety among spectra, with respect to given peaks, i.e. some peaks are like those in one or the other auto-spectrum, some are intermediate, some smaller than in either auto-spectra. In a sample of 64 cross-spectra from pairs of 72 auto-spectra, 14 cross-spectra have clear or strong peaks in the high-high frequency corner, compared to the auto-spectra where such peaks are much more common (34/72). Cross-spectra show low-low frequency peaks in 23/64, not much less commonly than in auto-spectra (29/72).
Cross-bicoherence can be asymmetrical around the equal frequency diagonal. The sample of 64 cross- spectra have 12 with obviously asymmetrical peaks and many more with less conspicuous ones. Most peaks are roughly symmetrical.
An exclusion "rule" is more commonly true than not, namely if there is a strong peak in the low-low frequency area, then there is no large peak in the high-high frequency area, and vice versa. This "rule" has 5 exceptions out of 64 in the foregoing sample. Auto-spectra show more common exceptions (14 / 72).
Brain states
Sleep & awake states. The statistics on all five subjects for whom suitable records and statistics are available (D, K, Ma, Me, R; >140 electrode loci) show a significant and consistent difference between waking and sleeping (stage II/III) states in the EEG samples of several minutes each - in all, 20 separate sleep episodes and nearly as many awake. In the sleep state, both in subdural and hippocampal loci, mean auto-bicoherence of all frequency pairs, epochs and electrodes ranges between 0.05 and 0.14, whereas in the waking state it is 0.03-0.10 and in each subject nearly always higher in sleep, on average by ca. 0.04. This is generally 2 or 3 times the 95% confidence intervals (Table III). It should be recalled that the mean value when there is virtually no significant bicoherence is ca. 0.04. Thus the greater value in sleep is relatively large. Evidence is suggestive that the fraction of nonlinearly coupled energy is highest in the EEG range <4 Hz in sleep whereas in the awake state the contribution from gamma band oscillations is more significant seizure state. The seizure state usually causes an increase in auto-bicoherence (Table IV) but there are many epochs after the "start of seizure" with no visible change in bicoherence, compared to the preceding, pre-ictal EEG. The distribution of high bicoherence is quite variable between seizures and electrodes, e.g. sometimes mainly in high frequencies, sometimes mainly in low.
|
|
Table III. Comparison of the awake, asleep and seizure states. Subject K; two episodes of each state; 16 subdural electrodes on frontal and parietal and anterior temporal lobes. Means (boldface) and 95% confidence intervals (small italics) for measures of the maximum power (maxp) in arbitrary units, maximum bispectrum (maxbis), maximum bicoh (maxbic) between zero and one, mean bicoherence (mnbic), skewness, (skew), asymmetry (asym) in the auto- and cross-spectra. From 6 files of ca. 2 min each, computed for 10 s epochs, pooling the values calculated for all epochs and for each of the 16 electrodes, except for the two rows of seizure measures from selected electrodes that show some stages of seizure activity during some of the epochs. Note that almost the only difference between the two episodes of the waking state and two of sleep is that mean bicoherence is higher in sleep. The ictal state in this sample shows distinctive characters only if the affected electrodes, judged from the raw record, are selected; they show principally an increase in bicoherence and asymmetry, besides the obvious increase in power and bispectrum. |
|
|
Table IV. Comparison of mean bicoherence in three brain states in several subjects. Means and 95% confidence intervals from many electrodes and 17 s epochs [except D = 9 s]. Sample files from each subject. All electrodes are pooled except in seizure files marked with * - in which electrodes are selected that show seizure activity has affected that locus. |
No one pattern of distribution of bicoherence over the bifrequency plane is consistent among the 20 seizure episodes in 11 subjects analyzed (Figs. 3, 4). Often there is a wide scatter of small peaks; sometimes there is a narrow mountain range parallel to one of the axes (like the ridge mentioned above). Some seizure files show a protracted period without spread of the electrical seizure beyond a few, or even one channel and the other loci may be subnormal in EEG power, with little or no bicoherence. Some stages of seizure activity in some seizures show little bicoherence. Even during conspicuous epochs of 4 Hz spike and dome waves, we observe in some files that mean bicoherence can be very low. Some seizures, which happen to be preceded by unusually high preictal bicoherence, show an actual overall reduction of bicoherence during the seizure. If we look more closely at the distribution of peaks over the bifrequency plane, there is no typical signature of seizures or of classical stages such as the spike and dome pattern. Of the several measures provided by the bispectral analysis, mean bicoherence is not always the one most clearly showing the change from the preictal to the ictal EEG. Sometimes maximum bicoherence, wherever it occurs in the bifrequency plane, or maximum bispectrum or skewness or asymmetry or total power shows the change more clearly (Fig. 8).
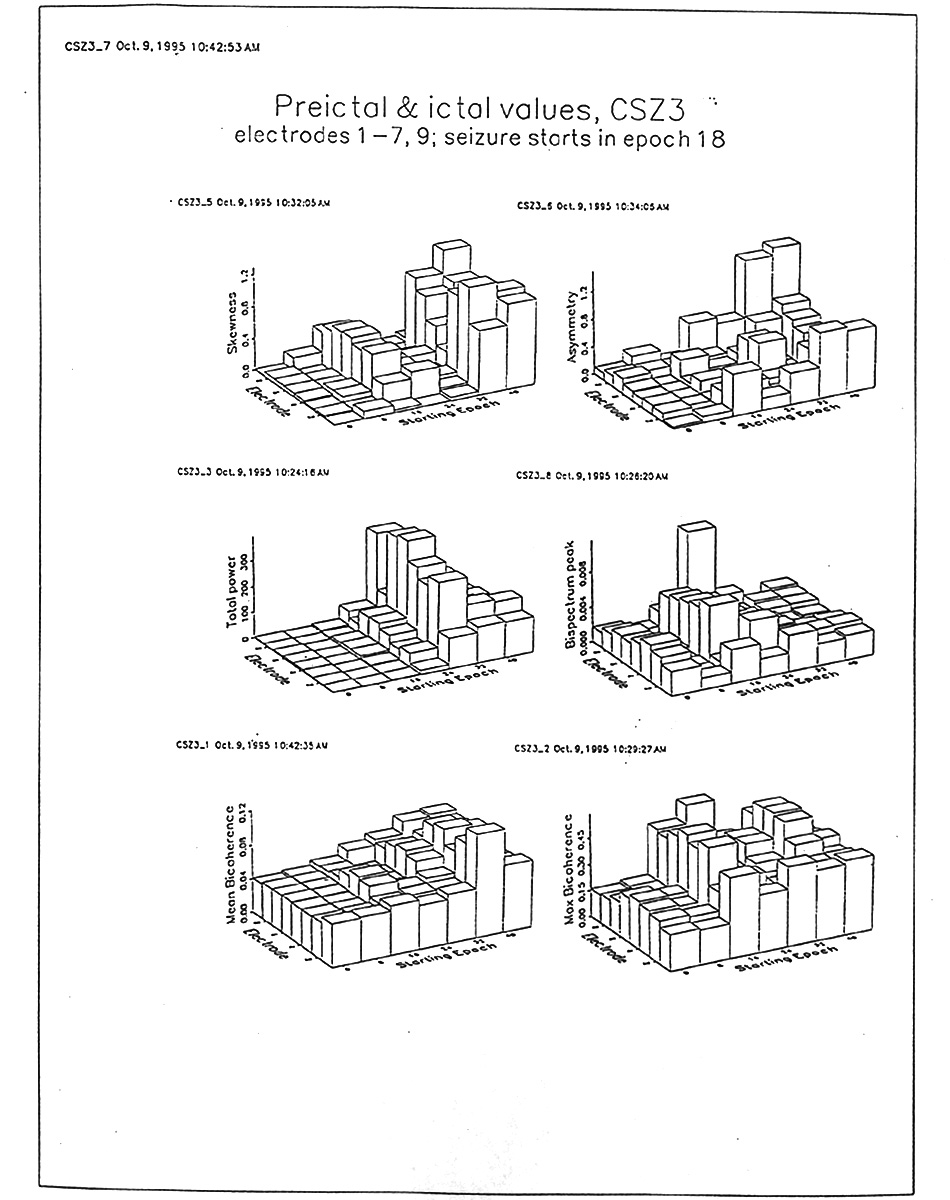 |
Figure 8. Comparison of several measures in bar graphs. Means from 8 electrodes for successive 9 s epochs of the ca. 60 s sample of EEG; first two columns of bars are pre-ictal; seizure starts with third column. Subject C, seizure 3. Skewness is averaged absolute magnitude, discarding sign. Each measure has disadvantages; e.g. maximum auto-bicoherence undervalues wide, low hills; mean auto-bicoherence undervalues narrow peaks. Note all these measures can jump abruptly from one electrode to the next and from one epoch to the next. |
Among three of the deep temporal lobe subjects from whom we have three seizures each, the seizures are quite diverse between the subjects and somewhat less so between the episodes in one subject. They progress through stages of distinct character. Some stages may never involve some electrode loci. Subject C is a case where the seizure is accompanied by substantial increases in maximum and mean auto- bicoherence, maximum and mean cross-bicoherence, skewness and asymmetry besides large increases in maximum and total power. Subject D, however, shows no average difference from the 6 min of preictal bicoherence to the 2'20" of seizure state and the 4'30" immediately after the seizure. Overall, the mean bicoherence of all electrodes and epochs during seizures ranges among subjects between 0.07 and 0.14, usually no higher than in slow wave sleep in the same subjects. If only those electrodes are chosen that show clear seizure activity, the mean bicoherence is hardly any or no greater than the mean of all electrode loci.
Most of the nine seizure starts in Subjects C, H and W, as determined by inspection of the raw EEG (V/t), are presaged by small but distinct increases in bicoherence, usually scattered or in the low-low corner of the bifrequency plane. This grows in successive 9s samples to large mountain ranges of bicoherence over many or all intersections (pairs of frequencies), except, usually, the high-high corner. That corner is occupied in the last epochs of the second seizure in Subject C (Csz2, 25+ seconds after seizure start), and in earlier epochs in the third seizure (Csz3 ca. 10 s after start.)
In some seizures (3 out of the nine in C, H and W) a conspicuous "bed of nails" (Figs. 3, 4) or hair- brush pattern of narrow bicoherence peaks in the low-low, mid-low and high-low regions coincides with quite regular and asymmetrical spikes at 3-5 Hz in the EEG and sharp teeth in the power spectra and a hair-brush in the 3D bispectrum. This means there are deep valleys of low bispectrum and bicoherence between the peaks at intersections of harmonics of 3-5 Hz up to at least the 6th or 7th harmonic. The impression is that when the pattern is a rounded mountain range or with fewer, irregular peaks and shallow valleys, i.e. a distribution of bicoherence over all frequency pairs, including nonharmonic ones, it is simply due to less regular intervals between asymmetrical EEG waves.
In seizures as in interictal states, neighboring electrodes can have extremely different bicoherence (6- 10 mm apart, center-to-center; each contact is quite large, being 1 mm wide bands around a 1 mm shaft), e.g. one electrode may show a high mountain or tall hair-brush and an adjacent one no bicoherence. This difference can sometimes correspond to a difference in the EEG between high amplitude paroxysmal spikes in one channel and normal background activity in the next. But quite often we can not see anything that would explain the observed difference in bicoherence, between different channels or adjacent epochs.
Brain regions.
In order to look for possible differences between loci or larger brain regions, such as the lobes, which were sampled by eight or ten electrodes each, cluster analyses were used combining three measures: mean bicoherence, asymmetry and skewness. As indicated above, these can vary independently although there is some average correlation. Fig. 9 is an example, comparing two regions of the same lobe, each represented by 8 electrodes and 4 epochs of 17 s each, i.e. 32 values. Such cluster plots show that there can be much overlap but also substantial differences between regions and lobes, given enough loci and time samples. We cannot, however, make any general statement about the consistency of such differences with our limited data set. The method remains an available, untried means to distinguish right and left, and cortical lobes. In the single subject with both subdural and deep temporal electrodes, the averages of many deep temporal loci are consistently different from those of many subdural cortical loci, in all three states: in the hippocampal loci mean and maximum bicoherence are always higher, on the average, but none of the other measures is consistently different.
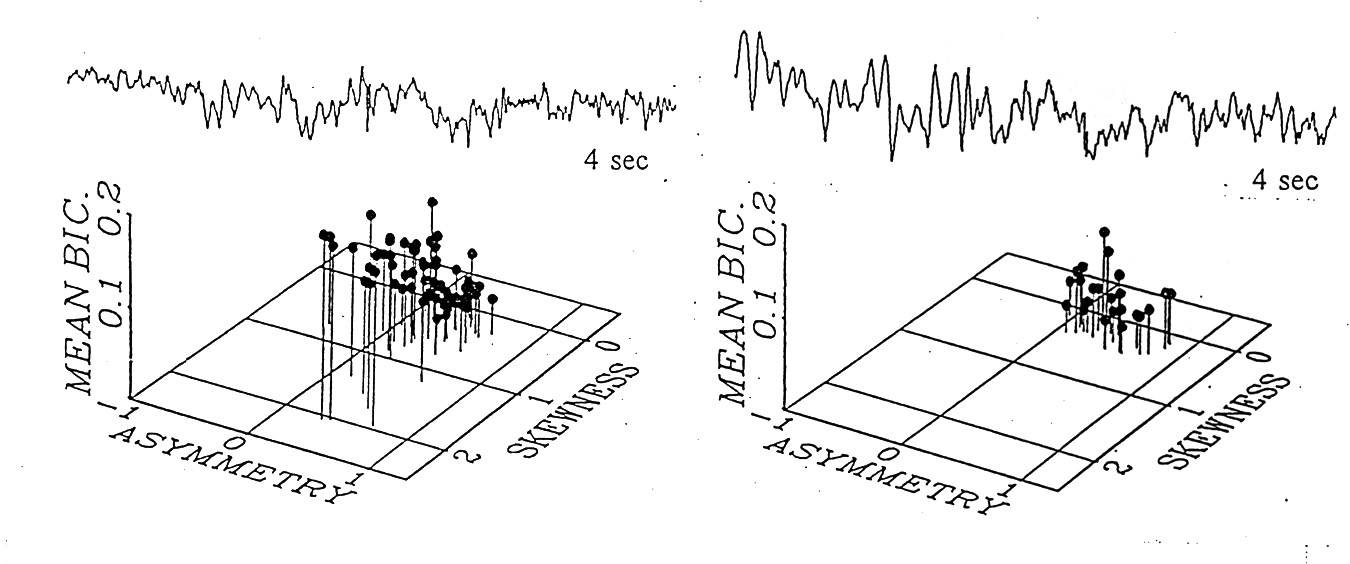 |
Figure 9. Comparison of two brain regions in the same subject at the same time, in an awake, interictal state, by cluster analysis; . Left, eight right subfrontal electrodes; right, eight right lateral frontal electrodes; four 17 s epochs. Each point represents the mean bicoherence over all intersections for one electrode for one epoch. Above, sample EEG from one of the loci in each cluster. Such cluster differences can be found between brain regions but are not yet known to be consistently characteristic. |
|
|
Discussion |
|
|
|---|
The most important conclusion from these findings is that a great deal more spatiotemporal information is carried in the intracranial EEG than the eye or familiar analyses can see. In many subdural and depth samples of 1.6 to 17 seconds we find episodically a form of cooperativity measured by quadratic phase coupling between pairs of frequency components, in the same or in different channels. Though commonly absent or nearly so, it is now and then prominent. Abrupt changes occur in the space of a few millimeters and a time span of one or two seconds. These numbers are based on our data set, which employed gross electrodes. It seems likely that the real dimensions, if we used semimicroelectrodes within the tissue, would be found to lie within tens of micrometers and milliseconds. Since different pairs of frequency components can be different in bicoherence and each can fluctuate rapidly, the picture resembles the bouncing light bars of an audio-amplifier frequency equalizer plus spatial differentiation in three dimensions.
Some EEG features like asymmetry and skewness clearly contribute to bicoherence, but most the features responsible for local or episodic bicoherence are usually not recognizable by visual inspection of the EEG, even when bicoherence is prominent. Highly similar looking segments of EEG a few seconds or a few millimeters apart may have either no appreciable auto-bicoherence or a substantial proportion of it. When present, it can be distributed in the bifrequency plane in any of a wide variety of patterns: periodic or aperiodic, solitary peaks like obelisks or broad mountain ranges, located in one or another or several parts of the plane, e.g. the high-high frequency region or high-low or low and intermediate or any other region. It is important to say that narrow, sharp peaks are not usual; relatively wide hills or mountains, including many contiguous frequency pairs are more common. Likewise, peaks at harmonic and subharmonic intersections are neither the rule nor particularly common.
Once accustomed to the idea that another and unfamiliar variable belongs within the scope of EEG dynamics, the important finding becomes its local, patchy distribution, fluctuating temporal status and complex 3-dimensional pattern on the bifrequency plane at each place and time. The patterns of these variables may turn out to characterize brain structures, ontogenetic stages, physiological or cognitive states or phylogenetic levels of complexity. Such characterization will not come easily, both because of the wide variety of wave forms that cause quadratic phase coupling and the high variance in time and space. The limited number of samples examined so far mainly emphasize great variability within a given structure and state. The evidence, in short, is in agreement with the original hypothesis that human cortical EEG is generally highly differentiated in respect to nonlinear cooperative interactions. This emphasis is the same as that reached in a separate study of the distribution of linear coherence between loci as well as successive time segments (Bullock and McClune, 1989; Bullock et al., 1995a, b), a measure quite independent of the bicoherence between frequencies. This emphasis on the local differentiation of what has been called the microEEG is not in conflict at all with the findings from scalp recording where a macrostructure exists that we could not predict from the sampling of microstructure.
We take the existence of bicoherence (defined by the fraction of non-Gaussian energy having a particular phase relationship among three frequencies: any two, F1, F2, and their sum, F3, such that the phase of F3 = the phases of F1 + F2) to be a sign of nonlinear cooperativity among frequency components of the compound field potential that arises from many generators. Whether the information carried in the cooperativity is itself a code (a signal and therefore influential in brain functioning) or is only a sign of the interactions among cellular and subcellular generators and their dynamic fluctuations (Bullock, 1997), in either case it becomes another descriptor potentially able to distinguish regions, stages, states and levels of brain evolution. As a quantitative and sensitive descriptor it might contribute to the understanding of what goes on in the brain, the more so as methods develop to evaluate different causes of bicoherence, due to transients or to ongoing relations such as special forms of amplitude or frequency modulation. Research on methods of estimating the relative contributions to the quadratic phase coupling of different kinds of ongoing wave forms (e.g. skewed or frequency modulated) and transient events (as in spikes or steps), is under way but still on the cutting edge (S. Elgar, pers. comm.). The abrupt changes in bicoherence here reported and the significant narrow peaks and valleys in plots of bicoherence vs time suggest that a common cause of quadratic phase coupling is episodic transients such as sharp corners and that this can be more important in some cases than steady state nonlinearity such as AM or ongoing asymmetry. Simulations have shown us that even a 10 ms pulse irregularly appearing about once per second causes widespread peaks detectable when the rms noise is as much as 0.2 the peak of the pulse. Transients can be expected to contribute all the more to bicoherence if events of the same shape are repeated, even if irregularly, as opposed to the same number of events of diverse shape.
We are not surprised to find bicoherence in the EEG, knowing that many departures from Gaussianity occur in the EEG. Several authors have already noted one or more peaks of bicoherence in small samples of EEG (cited in the Introduction). We were not prepared, however, for the extensive periods and places where no frequency pairs show any significant bicoherence or for the extreme diversity of the distribution of elevations and patterns of them all over the bifrequency plane even in loci a few millimeters apart, or for the rapid changes with time. We are surprised by the broad mountains as opposed to narrow peaks. We could not anticipate the curious forms of cross-bicoherence. We expected the finding of generally higher bicoherence during seizures but did not expect to find more bicoherence in sleep than in wakefulness. The state labelled wakefulness is probably heterogeneous. Our subjects were recorded on the ward under somewhat more natural living conditions than the standard EEG recording conditions. They almost never show periods of obvious alpha or other rhythms. We had no control of the open or closed state of the eyes or of attention, conversation or cognitive state and hence should not be surprised by the abrupt changes over time and the high variability.
Variability of bicoherence can be due to change in the absolute amount of phase coupling and also to change in the relative amount of non-quadratically coupled power at the corresponding frequencies. Thus some EEG segments with obvious skewness or numerous spikes may show low bicoherence from dilution with uncoupled activity or even phase-coupled activity of higher moments. Power line (60 Hz) interference should not affect our bicoherence values because we limit our view to the band below 50 Hz but muscle or other forms of extraneous spikes surviving the 100 Hz lowpass filter could either add spurious bicoherence or dilute the fraction of neural bicoherence, depending on their wave forms. If non- neural sources are adding spurious bicoherence in our data, they must be doing so in improbably short episodes. We have visually inspected the raw recordings and deleted the occasional bad channel; it is unlikely that we have missed sustained periods of artifacts. We do believe that some of the periods or channels of surprisingly low or insignificant bicoherence throughout the bifrequency plane, compared to the strong bicoherence in neighboring channels with similar and obviously skewed, asymmetric or spiky wave forms, may be due to dilution by a visible, sustained, low amplitude "fuzz" in the low bicoherence channels that, if it is responsible, must be wideband as well as symmetrical and unskewed.
An important finding is the independence of the variables: maximum peak power, total power (these are highly but not invariably or necessarily correlated in our samples), maximum bispectrum, maximum and mean bicoherence (usually rather highly correlated in our samples but of course either could be high when the other is low), skewness and asymmetry - each in the auto- and in the cross-spectra. Particularly interesting and to some, surprising, is the independence of bicoherence and both skewness and asymmetry (asymmetry of wave form around the time axis and the voltage axis, respectively), although the correlations are moderately high. Skewness and asymmetry are remarkably independent and poorly correlated with each other in our material.
In spite of the considerable number of electrode sites, epochs, and subjects, our material is insufficient to claim consistent differences between lobes or regions of the brain even within this set of individuals. The method may prove useful in some subjects to establish differences between homologous sites on the two sides or anomalies in particular lobes. As a diagnostic sign of seizure activity, bicoherence appears to be not as good as inspection of the raw EEG by a trained eye. As a candidate for automatic detection of seizures, it is doubtfully any better than simply measuring the total power in the power spectrum. It may, however, be a helpful complement. The typical differences reported between sleeping, waking and seizure states are at the moment less diagnostic than naked eye or linear spectral analyses, mainly because of the episodic nature of bicoherence amid long stretches of none.
As yet unexamined aspects might turn out to be the most significant features of bicoherence and especially of the bispectrum which we have not yet attempted to characterize systematically. These unexamined aspects include statistical comparison of the auto-bicoherence in different brain regions and more controlled brain states in a larger number of subjects, and of the several other measures of cross- bicoherence (e.g. subtractive, additive, F1 and F2 from different channels) for the same regions and states. Also in this list are the possibility of mapping loci with more cross-bicoherence than the general average, any rules about the function of distance between loci, the frequency-specific distribution of quadratic phase coupling and of skewness and asymmetry with brain state or region, and the degree of independence of local coherence and bicoherence. Our brain states are not well controlled and leave open the more subtle cognitive states. We have not compared the scalp EEG with the intracranial; suitable data is rare for technical reasons.
This close look at EEG bicoherence, besides the surprises mentioned above, has compelled us to press the mathematical experts for help in dissecting the relative roles of the known sources of quadratic phase coupling. These can apparently be subsumed under two rubrics, with subdivisions. (i) Ongoing rhythms that have modulations of amplitude, frequency or phase by other rhythms can cause bicoherence, but only if some of the modulating signal is present in addition to the modulated mixture (note the experiment described under Methods, with artificial data). Modulations also include those distortions of sine waves that are either sawtooth (or otherwise asymmetrical around the voltage axis) or scalloped (rounded peaks and sharp valleys: skewed around the time axis). (ii) Transient events with more or less sharp corners at their onset, offset or change in slope introduce a wide band of time-locked, phase-coupled frequencies. Although we have numerical estimates of the skewness and asymmetry, we do not know their relative contributions to the bicoherence compared to other causes. It will be valuable to learn how to evaluate the classes of contributions.
In the meantime we can speculate that transient events may play a larger role in the dynamics and cooperative properties of EEG than has been recognized heretofore. These considerations also serve to emphasize that bicoherence is not a measure of a simple or a single operation or form of interaction but a mathematical relation among the artificially segregated frequency components of one or two time series that can come from a variety of wave forms, continuous or transient. Interpretation of what it means for brain operations when bicoherence suddenly appears, in one or another pattern on the bifrequency plane, is therefore not to be expected, pending new tools. What it means for brain understanding is two-fold. (i) It can direct attention to certain episodes and loci, certain wave forms and frequency bands that might yield to closer inspection. (ii) It provides a powerful descriptor, not predictable from familiar analyses, that might be found to distinguish certain states, stages, regions or taxa.
|
|
Acknowledgement/References |
|
|
|---|
Acknowledgement This work was aided by grants from the National Institute of Neurological Diseases and Stroke. Robert Guza and Stephen Elgar have provided crucial expert advise and a number of friends have helped with the manuscript.
Achimowicz, J.Z. and Bullock TH. Nonlinear properties of local field potentials in brain: implications for biological neural networks modelling. Proc. Annu. Res. Symp., Inst. Neural Computation, UCSD, 1993a, 3: 29-49.
Achimowicz, J.Z. and Bullock, T.H. Nonlinear properties of EEG: bispectrum in human and animal brains. Soc. Neurosci. Abstr., 1993b, 19: 1605.
Barnett, T.P., Johnson, L.C., Naitoh, P., Hicks, N. and Nute, C. Bispectrum analysis of electroencephalogram signals during waking and sleeping. Science, 1971, 172: 401-402.
Bullock, T.H. The micro-EEG represents varied degrees of cooperativity among wide-band generators: spatial and temporal microstructure of field potentials. In: E. Basar and T.H. Bullock (Eds.). Brain Dynamics: Progress and Perspectives. Springer-Verlag, Berlin, 1989: 5-12.
Bullock, T.H. Signals and signs in the nervous system. Proc. Nat. Acad. Sci. (USA), 1997, 94:1-6.
Bullock, T.H. and Achimowicz, J.Z. A comparative survey of event related brain oscillations. In: C. Pantev, T. Elbert, B. Lütkenhöner (Eds), Oscillatory Event-Related Brain Dynamics, NATO ASI Series, Vol. 271. Plenum Press, New York, 1994, 11-26, Figs. 8, 9.
Bullock, T.H. and McClune, M.C. Lateral coherence of the electrocorticogram: a new measure of brain synchrony. Electroencephalog. clin . Neurophysiol., 1989, 73: 479-498.
Bullock, T.H., McClune, M.C., Achimowicz, J.Z., Iragui-Madoz, V.J., Duckrow, R.B. and Spencer, S.S. Temporal fluctuations in coherence of brain waves. Proc. Nat. Acad. Sci. (USA), 1995a, 92: 11568- 11572.
Bullock, T.H., McClune, M.C., Achimowicz, J.Z., Iragui-Madoz, V.J., Duckrow, R.B. and Spencer, S.S. EEG coherence has structure in the millimeter domain: subdural and hippocampal recordings from epileptic patients. Electroencephalog. clin. Neurophysiol., 1995b, 95: 161-177.
Chandran, V. and Elgar, S.L. Pattern recognition using invariants defined from higher order spectra - one dimensional inputs. IEEE Trans. on Signal Processing, 1993, 41: 205-212.
Dumermuth, G. and Gasser, T. The computation of EEG bispectra. Comput. Prog. Biomed., 1978, 8: 235-242.
Dumermuth, G. and Molinari, L. Spectral analysis of EEG background activity. In: A.S. Gevins and Rémond, A. (Eds.), Methods of Analysis of Brain Electrical and Magnetic Signals. EEG Handbook, revised series, Vol. 1. Elsevier Science Publishers B.V., Amsterdam, 1987: 85-130.
Dumermuth, G., Huber, P.J., Kleiner, B and Gasser, T. Analysis of the interrelations between frequency bands of the EEG by means of the bispectrum. A preliminary study. Electroencephalog. clin. Neurophysiol., 1971, 31: 137-148.
Dumermuth, G., Gasser, T. and Lange, B. Aspects of EEG analysis in the frequency domain. In: D. Dolce and H. Kuenkel (Eds.), Computerized EEG analysis (CEAN). Gustav Fischer, Stuttgart, 1975, 432- 457.
Dumermuth, G., Lange, B. and Herdan, M. Analyse spectrale de l'activite EEG rapide (beta). Rev. Electroencephalog. Neurophysiol. Clin., 1983, 13: 122-128.
Eckhorn, R. Oscillatory and non-oscillatory synchronizations in the visual cortex of cat and monkey. In: C. Pantev, T. Elbert, B. Lütkenhöner (Eds.), Oscillatory Event-Related Brain Dynamics, NATO ASI Series, Vol. 271. New York, Plenum Press, 1994: 115-134, Fig. 17.
Elgar, S. and Guza, R.T. Observations of bispectra of shoaling surface gravity waves. J. Fluid Mech., 1985, 161: 425-448.
Elgar, S. and Guza, R.T. Statistics of bicoherence. IEEE Trans. Acoust. Speech and Sign Proc., 1988, 36: 1667-1668.
Elgar, S. and Sebert, G. Statistics of bicoherence and biphase. J. Geophys. Res., 1989, 94: 10,993-10,998.
Freeman, W.J. Predictions on neocortical dynamics derived from studies in paleocortex. In: E. Basar and T.H. Bullock (Eds.), Induced Rhythms of the Brain. Birkhäuser Boston, Cambridge, MA, 1992: 183- 199.
Freeman, W.J. and Baird, B. Relation of olfactory EEG to behavior: spatial analysis. Behav. Neurosci., 1987, 101: 393-408.
Freeman, W.J. and Davis, G.W. Olfactory EEG changes under serial discrimination of odorants by rabbits. In: D. Schild (Ed.), Chemosensory Information Processing. NATO ASI Series H39. Springer- Verlag, Berlin, 1990: 375-391.
Freeman, W.J. and Skarda, C. Spatial EEG patterns, nonlinear dynamics and perception: the neo- Sherringtonian view. Brain Res. Rev., 1985, 10: 147-175.
Freeman, W.J. and Vianna di Prisco, G. Relation of olfactory EEG to behavior: time series analysis. Behav. Neurosci., 1986, 100: 753-763.
Hasselman, K., Munk, W. and MacDonald, G. Bispectra of ocean waves. In: M. Rosenblatt (Ed.), Time Series Analysis. John Wiley, New York, 1963, 125-139.
Kearse, L.A.J., Manberg, P., DeBros, F., Chamoun, N. and Sinai, V. Bispectral analysis of the electroencephalogram during induction of anesthesia may predict hemodynamic responses to laryngoscopy and intubation. Electroencephalogr Clin Neurophysiol., 1994, 90: 194-200.
Kim, Y.C. and Powers, E.J. Digital bispectral analysis and its applications to nonlinear wave interactions. IEEE Trans. on Plasma Science, 1979, PS-7: 120-131.
Lopes da Silva, F.H. Dynamics of EEGs as signals of neuronal populations: models and theoretical considerations. In: E. Niedermeyer and F. Lopes da Silva (Eds.), Electroencephalography: Basic Principles, Clinical Applications and Related Fields. Urban and Schwarzenberg, Baltimore-Munich, 1987: 15-28.
Lopes da Silva, F.H. and Storm Van Leeuwen, W. The cortical alpha rhythm in dog: the depth and surface profile of phase. In: M.A.B. Brazier and H. Petsche (Eds.) Architectonics of the Cerebral Cortex. Raven Press, New York, 1978: 319-333.
Mendel, J.M. Tutorial on higher-order statistics (spectra) in signal processing and system theory: theoretical results and some applications. Proc.IEEE, 1991, 79: 278-305.
Nikias, C.L. and Raghuveer, M.R. Bispectrum estimation: a digital signal processing framework. Proc. IEEE, 1987, 75:867-891.
Ning, T. and Bronzino, J.D. Bispectral analysis of the rat EEG during various vigilance states. IEEE Trans. Biomed. Eng., 1989, 36: 497-499.
Pezeshki, C., Elgar, S. and Krishna, R.C. Bispectral analysis of systems possessing chaotic motion. J. Sound Vib., 1990, 137: 357-368.
Samar, V.J., Swartz, K.O., Raghuveer, M.R., Braveman, A.L., Centola, J.M. and Jackson J.K. Quadratic phase coupling in auditory evoked potentials from healthy old subjects and subjects with Alzheimer's dementia. In: A.Giannakis (Ed.), IEEE Signal Processing Workshop on Higher Order Statistics, South Lake Tahoe, 1993: 361-365.
Sherman, D.L. Novel Techniques for the Detection and Estimation of Three-Wave Coupling with Application to Human Brain Waves. Ph.D. Thesis, Purdue University, 1993.
Sherman, D.L. and Zoltowski, M.D. Application of eigenstructure based bispectrum estimation: EEG wave coupling in cognitive tasks. In: (Anonymous ed.) Workshop on Higher-Order Spectral Analysis, Vail, Colorado, Sponsored by Office of Naval Research and NSF, 1989, 135-139.
Sigl, J.C. and Chamoun, N.G. An introduction to bispectral analysis for the electroencephalogram. J. Clin. Monit., 1994, 10: 392-404.
Tang, Y. and Norcia, A.M. Coherent bispectral analysis of the steady-state VEP. Proc. IEEE Eng. Med. Biol. Soc., 1996, 17: 22-24.
Whitton , J.L., Elgie, S.M., Kugel, H. and Moldofsky , H. Genetic dependence of the electroencephalogram bispectrum. Electroencephalog. clin. Neurophysiol., 1985, 60: 293-298.