T.H. Bullock(1), M.C. McClune, J.Z. Achimowicz, V.J. Iragui-Madoz, R.B. Duckrow†and S.S. Spencer*
Department of Neurosciences, University of California, San Diego, La Jolla, CA 92093-0201
† Department of Neurology, University of Connecticut Health Center, Farmington,CT 06030
*Department of Neurology, Yale University School of Medicine, New Haven CT 06510
Corresponding author:
T.H. Bullock
Tel. (619) 534-3636. Fax: (619) 534-3919;
E-mail: tbullock@ucsd.edu
Key words: Sleep; Seizure; Depth recording; Hippocampus; Cooperativity
|
|
Summary |
|
|
|---|
Subdural recordings from 8 patients via rows of eight electrodes with either 5 or 10 mm spacing plus depth recordings from 3 patients with rows of 8-12 electrodes either 6.5 or 9 mm center-to-center were searched for signs of significant local differentiation of coherence calculated between all possible pairs of loci. EEG samples of 2-4 min were taken during four states: alertness, stage 2-3 sleep, light surgical anesthesia permitting the patient to respond to questions or commands, and electrical seizures. Coherence was computed for all frequencies from 1-50 Hz or 0.3-100 Hz and then compared for 6 or 7 narrower bands between 2 and 70 Hz.
In both the subdural surface samples and those from temporal lobe depth electrode arrays coherence declines with distance between electrodes of the pair, on the average. This is nearly the same for all frequency bands. Whether computed for 5, 20 or 60 s epochs, coherence pooled across all pairs of a given separation, in a given subject, differs only slightly, in the direction of lower coherence for longer samples, indicating good stationarity of the samples chosen. For middle bands like 8-13 and 13-20 Hz, mean coherence typically declines most steeply in the first 10 mm, from values indistinguishable from 1.0 at <0.5 mm distance to 0.5 at 5-10 mm and to 0.25 in another 10-20 mm in the subdural surface data. Temporal lobe depth estimates decline ca. half as fast; coherence 0.5 extends for 9-20 mm and 0.25 for another 20-35 mm. Low frequency bands (1-5, 5-8 Hz) usually fall slightly more slowly than high frequency bands (20-35, 35-50 Hz) but the difference is small and variance large. The steepness of decline with distance in humans is significantly but only slightly smaller than that we reported earlier for the rabbit and rat, averaging < one half.
Local coherence, for individual pairs of loci, shows differentiation in the millimeter range, i.e. nearest neighbor pairs may be locally well above or below average and this is sustained over minutes. Local highs and lows tend to be similar for widely different frequency bands. Coherence varies quite independently of power, although they are sometimes correlated.
Regional differentiation is statistically significant in average coherence among pairs of loci on temporal vs frontal cortex or lateral frontal vs subfrontal strips in the same patient, but such differences are usually small. We could not test how consistent they are over hours or between patients. Differences between left and right hemispheres, whether symmetrical pairs or pooled from two or more lobes on each side, can be quite large; in our patients the right side is usually higher, especially in the waking state.
Brain state has a large influence. Slow wave sleep usually shows slightly more coherence at each distance, in all bands, compared to the waking EEG, but not consistently. Coherence at a given distance or its rate of decline with distance is a more direct measure of synchrony than naked-eye "synchronization," which is dominated by the power spectrum. Among the range of EEG states classified as seizures, coherence varies widely but averages higher by 0.05-0.2 than in pre-ictal states, usually in all frequencies when computed over the whole seizure but much more in the higher bands during the height of the electrical paroxysm.
The findings point to still finer structure and more variance with closer spacing of electrodes. They could not predict the known large scale coherence between scalp electrodes, but are not in conflict with them. Scalp recording blurs the finer spatial structure, but reveals macrostructure missed by subdural and depth recording with limited numbers of channels. The strong tendency for correlated fluctuations across frequency bands is contrary to expectation from the common model of independent oscillators.
|
|
Introduction |
|
|
|---|
Previous work on EEG coherence in rats and rabbits (Bullock and McClune 1989) showed that coherence between pairs of electrodes on the pial surface falls with distance from the maximal value of 1.0, which it must have at a small enough distance in a volume conductor so that the cones of tissue seen by the two electrodes overlap virtually completely, to values around 0.5 at ca. 3-6 mm, when averages were computed over tens of seconds of EEG from many electrode pairs for each distance. This rather steep decline, and the wide variance between pairs, suggested that there might be structure in the millimeter domain. The next step to be taken was to look at individual pairs of electrodes, instead of pooling all pairs with the same separation, to discover whether there are signs of local differentiation, whether they are stable over at least several seconds and whether they are characteristic of cortical areas or states. The opportunity to do this in human data arises from the availability of EEG records recorded with arrays of subdural and depth electrodes routinely implanted in patients with intractable epilepsy (Engel et al. 1990; Mackenzie et al. 1990; Mihara et al. 1992; Duckrow and Spencer 1992; Garcia de Leon 1993; Kanner et al. 1993). Signs of fine structure in the distribution of coherence are not expected in the usual scalp records of the human EEG, in which each electrode sees a large cone of brain tissue.
The aim of this study is to explore the spatial distribution of this measure of neuronal cooperativity between pairs of recording sites on and in the human brain, on a relatively fine scale. The finding that there are local discontinuities in the millimeter domain has significance in underlining earlier indications that the view of EEG distribution from scalp recording can miss completely most of the structural organization of activity. It will be shown that not only is the finding of local fine structure typical of our sample of human EEGs, but in addition unexpected similarities of coherence distribution for widely different frequency bands are common. This must have profound consequences for models of the origin of the EEG. The relatively meager relevant literature is cited in the Discussion. Preliminary reports have appeared (Bullock et al. 1990, 1992).
|
|
Methods |
|
|
|---|
Data from the first 8 patients were supplied by V.J.I-M from the U.C.S.D. Epilepsy Center. Seven of these patients had been implanted by neurosurgeon, Dr. J.F. Alksne, with plastic subdural strips carrying rows of eight electrodes at 10 mm intervals, for long term recording, for localization of seizure foci. A given patient may have strips bilaterally on frontal, temporal and parietal lobes, the exact position and orientation varying among the patients (Fig. 1). Originally the EEG was recorded continuously for many days. We took 2-4 min samples during slow wave (stage 2-3) sleep, alert, interictal periods and during electrical seizures. Additional recordings were performed in the operating room on the exposed brain when the patient was in an alert, sedated state, able to respond to commands and questions (agents commonly used were nitrous oxide, Fentanyl and Forane). Such records are valuable because electrode grids with 5 mm spacing were used. An eighth patient gave data only from such operating room recording.
Depth recordings from 3 additional patients from the Yale University Epilepsy Surgery Program were made available by S.S.S. and R.B.D. These patients had been implanted by Dr. D.D. Spencer with medial temporal lobe 8-12 contact depth electrode arrays, placed under MRI control, bilaterally, in the parasagittal planes such that the most anterior electrodes were in the amygdala, the next in the pes hippocampi, several in the anterior hippocampus and the last few in posterior hippocampus, longitudinally (diagrams of the placement are given in Duckrow and Spencer 1992). Additional channels sampled overlying temporal cortex of the superior, middle or inferior gyrus or fusiform gyrus or entorhinal cortex of one or both sides, but were too few to contribute to the present analysis of distribution of coherence.
The actual electrode contacts in the subdural surface strips with 10 mm center to center spacing are metal disks, 2 mm in diameter. The metal disks in the grids with 5 mm spacing are slightly smaller. The contacts on the 1 mm diameter temporal lobe depth electrode arrays are nichrome collars 2 mm long x 1mm in diameter around a polyurethane tube, spaced either 6.5 or 9 mm center to center.
For the purposes of this study, bipolar recording was rejected as more inconsistent and difficult to interpret than monopolar recording with a well chosen, quiet, reference. The reference electrode for the subdural records was common, extracranial, and usually on linked ears; for the depth records it was common and either on one mastoid process or one ear. In view of the warnings in the literature that coherence with a common reference is liable to contamination by the activity of the common electrode (Fein et al. 1988; Rappelsberger 1989), it is important to point out that our reference electrodes must have been quite inactive as shown by the fact that very low coherence values were common. Any low value means that the shared activity in the two channels from the common electrode must have been very small since a common signal usually gives coherence well above the bias level even when it is a small fraction of the total activity, depending on its "color" or spectral distribution.
The subdural records were digitized at 240 Hz after first order (RC) filtering from 1-70 Hz. The depth records were digitized at 200 Hz after similar filtering from 0.3-100 Hz. Using the Fast Fourier Transform (FFT), the spectra for power, phase and coherence were computed and plotted for each possible pair of electrodes. Magnitude-squared coherence values were computed for each frequency and each pair of electrodes, in epochs of 1 s and averaged for relatively stationary EEG samples of 5, 20 or 60 s or 2 min. Coherence values were corrected for the "bias", the value expected for the case of minimal coherence, determined by coincidence from two quite independent sources; it depends on the duration of the sample, the digitizing rate and frequency band and assumes linearity and independence of frequencies. Coherence cannot have a negative value and only approaches zero in long samples of independent time series.
|
|
Results |
|
|
|---|
In the following sections on coherence with frequency, distance, individual loci, lobes and brain states we deal first with the distribution of coherence in arrays of electrodes indwelling for many days under the dura mater over parietal, temporal and frontal cortex. Then follow results from the depth electrode arrays in the medial temporal lobe in three patients for each of whom we have pre-ictal and ictal periods of three seizures.
Comparison of the coherence between pairs of loci in the depth recordings, mainly from the hippocampus, with the coherence values between pairs of subdural loci, mainly from parietal, temporal and frontal cortex, faces some asymmetries of unknown significance. The electrodes are of different form, so that more surface area is in contact with brain tissue in the depth electrodes than in the surface arrays. The spacing is 5 or 10 mm in the latter and 6.5 or 9 mm in the former. Still, the differences in mean coherence in the alert, interictal state are probably real and not only due to such asymmetries. The mean coherences for 6 bands pooling at least one minute of EEG from all 10 mm subdural pairs in 7 patients in this state range from 0.34-0.47; the means of three 20 s pre-seizure samples of all 9 mm pairs of deep temporal electrodes in the two subjects with that spacing range for the same bands from 0.48-0.61 (Table 1).
| Filter band, Hz | 2-5 | 5-8 | 8-13 | 13-20 | 20-35 | 35-50 |
| Subdural C10mm | 0.47 | 0.47 | 0.43 | 0.40 | 0.34 | 0.38 |
| Depth probe C9mm | 0.48 | 0.61 | 0.55 | 0.49 | 0.49 | 0.51 |
A. Coherence as function of frequency
In the subdural, surface data coherence, power and phase spectra were examined for each pair of electrodes. The distribution of coherence values among many pairs, each averaged over about 2 min of EEG, has been plotted for limited frequency bands in a number of samples in different brain states (Fig. 1). It often approximates a Gaussian distribution, without any obvious tendency to skewness or multimodality, but there are, of course, macro- and micro-states defined by fluctuations in these properties. Variance of coherence is usually between 4 and 20% of the mean. The mean depends, among other things, upon the frequency band, the state, and the distance between the two electrodes of the pair. These last two factors are treated in subsequent sections. Three findings about the dependence on frequency are of special interest.
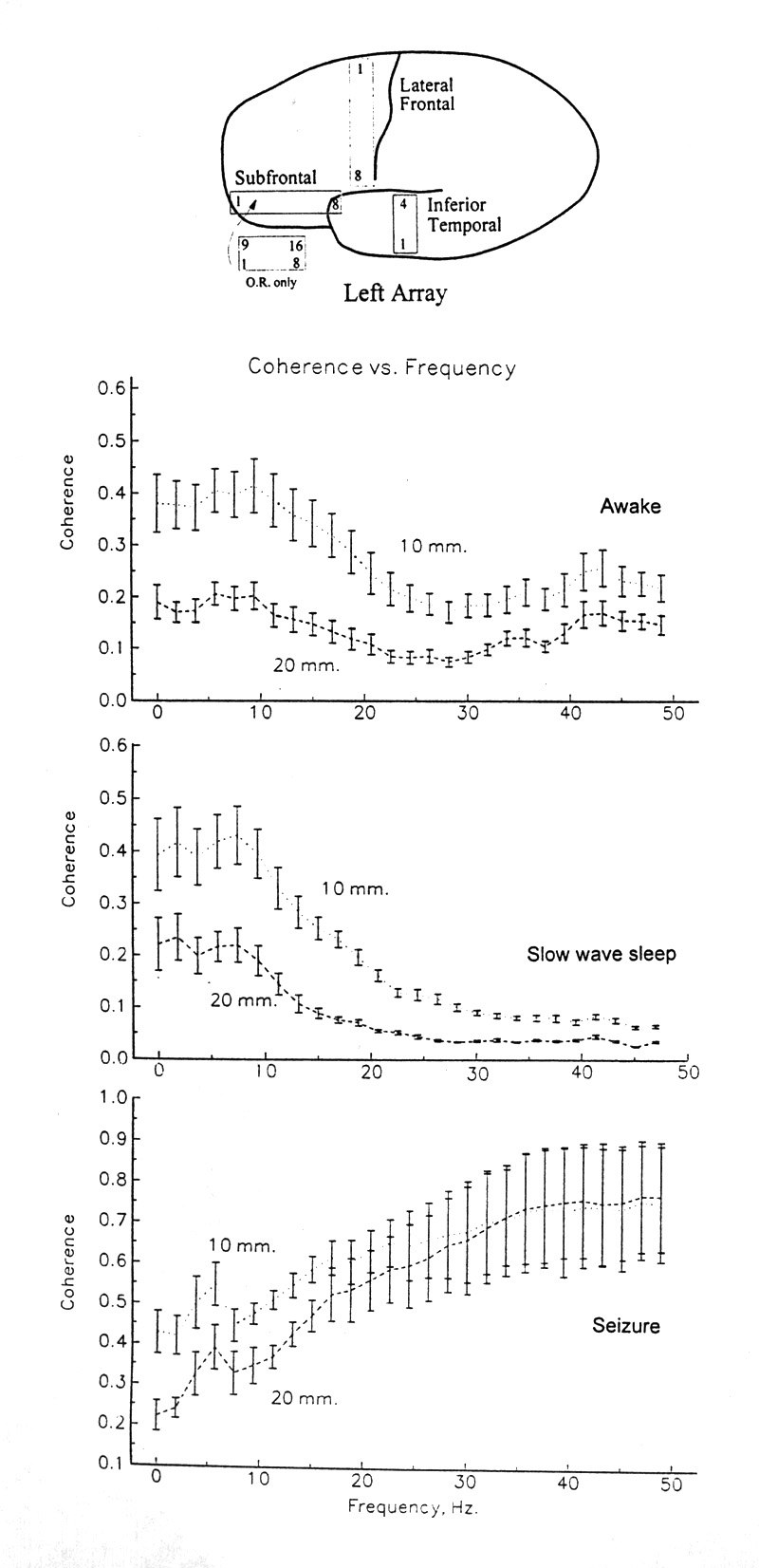 |
Figure 1. Coherence as function of frequency. Means and standard errors of the coherence values computed from 2 min of EEG from all pairs of electrodes 10 mm apart and all pairs 20 mm apart, from Patient 7 in three states, for each successive 20 s epoch, with a resolution of ca. two Herz. Top panel, awake; second panel, in slow wave sleep; third panel, during a seizure. Note the large variance with high coherence at high frequency during seizure; this is peculiar to these epochs. Variance in other samples decreases at high coherence. Separation of electrodes by only 10 mm reliably causes lower coherence. Bottom, cartoon of brain and position of subdural plastic strips in another case, Patient 6. The wide strip shown outside the brain had 2 rows of electrodes at 5 mm spacing and was used only in the operating room. All other strips had one row of electrodes at 10 mm spacing and were indwelling for some days. The numbers identify electrodes at the ends of rows. A symmetrical set of strips was placed on the right side. Operating room recording over the convexity was usually done with square, 35 x 35 mm arrays of 8 x 8 electrodes with 5 mm spacing. |
(i) Coherence commonly falls somewhat with increasing F, especially above our band 2 (5-8 Hz), but the decline is typically barely significant. Many spectra show little or no fall.
(ii) Many spectra show a marked upturn in coherence above 20 Hz and especially in the 35-50 Hz band (Fig. 1, top, Tables 1 and 3), often quite steep. This character does not act like the end of a continuous distribution from negative to positive slopes in the upper frequencies but more like an either/or, gentle down-slope or steep up-slope. In 18 mean spectra from 7 patients in both sleeping and waking states (Table 3), each mean representing 120 pairs, or close to it, and one of six arbitrary bands of frequencies and at least three 20 s epochs of EEG, seven of the 18 show the high frequency up-turn of coherence, all highly significant. This striking dichotomy does not correlate with the sleeping or waking state but occurs in a third to a half of the samples in both states. During seizure states it is usual that coherence rises in bands 5 and 6. This, however, is not invariable; the presence or absence of this character is one of several ways in which the seizure state appears to be heterogeneous in coherence.
(iii) Coherences for the bands up to 20-35 Hz tend to fluctuate together; those places or pairs with higher coherence typically have higher coherence in all these bands; those with low coherence typically are low in all bands. The rather common up-slope for bands 5 and 6 is the most conspicuous and almost the only exception to this rule. The same rule applies to fluctuations in coherence over time, to be dealt with in a separate paper.
In the temporal lobe depth recordings the principal result, as for subdural recordings, is that frequency has only a small influence. Coherence tends to be smaller as frequency increases in averages of larger samples; this dependence is typically continuous over our frequency range and rarely exceeds 33% of the highest value (lowest band). Without averaging, the relation is extremely variable and it is notable that in averages of smaller samples, when coherence is low in one band, it tends to be low in all bands. There is a significant non-independence between bands.
A common departure from the approximately monotonic coherence decline with frequency is an upturn in the bands above 30 Hz, sometimes beginning at 20 Hz, quite similar to that just noted for subdural recordings.
B. Coherence as function of distance between electrodes of a pair
Coherence in the human EEG quite consistently falls with distance, at least within the first 40-50 mm in subdural (Figs. 2-5) and in depth recordings (Fig. 6). At greater distances, such as between regions, lobes or hemispheres, it can level off or even rise. The steepness of decline in the first 40-50 mm varies, but not greatly. It approximates linearity on a semi-log plot (log distance) but is somewhat steeper at shorter distances. We can justify plotting to a coherence value of 1.0 at zero distance and then note that the slope tends to be steeper in the first 10 mm and in the higher frequency bands, except for the 35-50 Hz band where it comes back to a shallower slope. The mean subdural coherence for electrodes 10 mm apart (C10mm) is usually 0.4-0.5 in the awake, interictal state for EEG in frontal, temporal and parietal cortex. For comparison with other species, the typical distance for a coherence of 0.5 (D0.5) is 5-10 mm, subdurally, and 9-20 mm with depth electrodes in the region of the hippocampal formation.
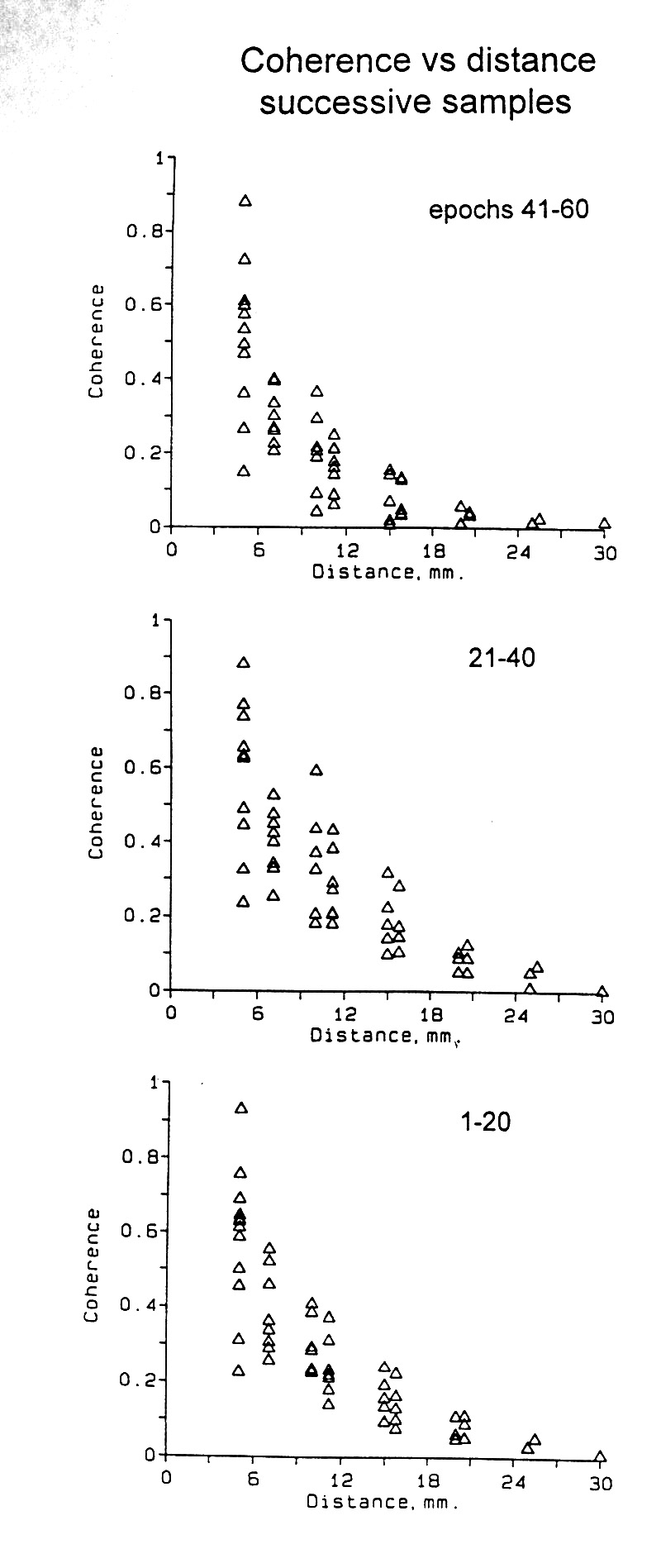 |
Figure 2. Coherence as function of distance. Scatter graph of individual values of coherence in the 8-13 Hz band for 20 s epochs for all electrode pairs up to 30 mm apart in an array with two adjacent rows of subdural electrodes over the right mid-frontal lobe of Patient K under sedation in the operating room. The panels show successive 20 s epochs to indicate the relatively high stationarity. The N of pairs is shown in each panel, with the same set of pairs and their decreasing number at the greater separations. |
A convenient measure of steepness, with our subdural data from electrodes usually spaced 10 mm apart, is the ratio C20mm/C10mm; this averages to 0.61 in a large pool of awake, interictal pairs from 6 patients (see Table 2 for ratios by frequency band); this means that coherence between pairs of electrodes 20 mm apart declines on average to 61% of the coherence of 10 mm pairs. There is a slight tendency to steeper decline in higher frequency bands, except for the highest (35-50 Hz) band, which falls only to the same extent as the lowest band. These numbers say that the decline in coherence with distance is a little steeper in the first 10 mm than in the second 10 mm.
 |
Figure 3. Coherence as function of length of epoch. EEG for two minutes from of patient K was divided into two 60 s or 24 5 s epochs for computing coherence. The close agreement indicates high stationarity and was typical, not only of the sedated, responsive state (upper panels) but of the alert state without drugs (lower panels), as well as the stage II sleeping state. During seizure, coherence tends to be lower in longer samples. Note the abscissae do not begin at 0 mm. A line can be justified beginning at coherence ca. 1.0 at a distance <0.1 mm, where the two electrodes essentially overlap completely in the cone of tissue they "see." |
The decline of coherence with distance is generally not significantly different for various frequency bands but a common tendency is for the highest band (35-50 Hz) to fall the most, especially at 20-30 mm or more.
| Filter band, Hz: | 2-5 | 5-8 | 8-13 | 13-20 | 20-35 | 35-50 |
| Ratio: C20mm/C10mm | 0.65 | 0.62 | 0.63 | 0.57 | 0.54 | 0.65 |
In the hippocampal electrodes, as on the subdural isocortical surface, coherence declines clearly and significantly with distance between the electrodes of a pair. Figure 6 shows the 20 s means of all pairs of the 8 contacts along the depth array, plotted by distance. Since a point can justifiably be added for coherence equal to 1.0 at a distance close to zero, we look at the distance where coherence is 0.5 and find it at <9 to 20 mm in almost all bands and samples; only the highest band (50-70 Hz) in one subject (H) fails to decline to 0.5 by 35 mm. Coherence is high at 6.5 mm and falls to about 1/2 of that by 20-35 mm in each of the four subjects and in all 7 bands, up to 70 Hz. After the initial steep decline, all subjects and bands show a slower fall or a plateau or an actual upturn above 35-40 mm. The upturn, like the plateau in subject H, is seen principally in the highest band. As in subdural cortical activity, the means represent a wide scatter of values for different epochs, pairs and the three samples taken prior to different seizure episodes; the scatter is reduced if the epoch used for computation is increased.
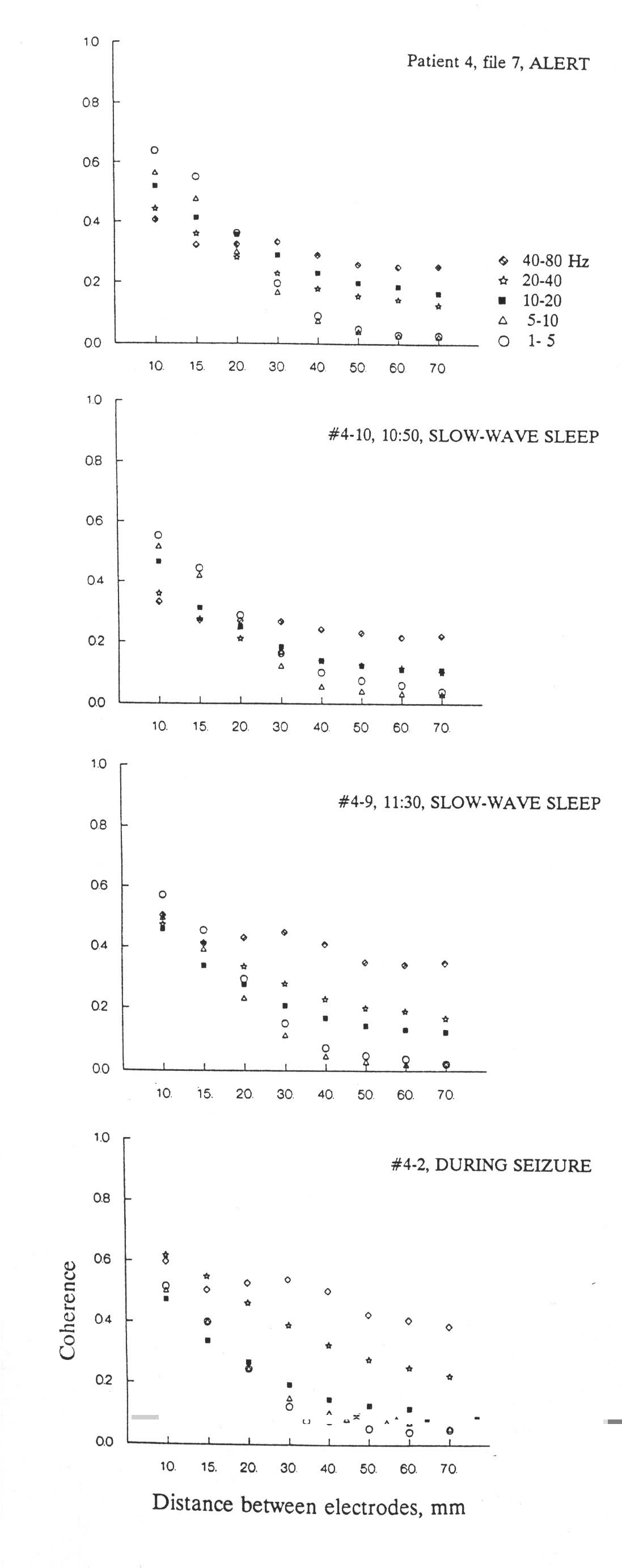 |
Figure 4. Coherence for several states. Subdural recording, Patient 4, parieto-occipital cortex; 16 electrode channels, 2 min of data computed for 60 s epochs for all 120 pairs. Mean values for the stated frequency bands for all pairs of the given distance. Two samples during stage II sleep show that there may be a significant enhancement of synchrony (#4-9, 11:30) in the highest band, compared with the alert state but at least as often there is no increase in synchrony in any band (#4-10, 10:50). Seizures typically increase synchrony at least in the higher bands and often also in lower bands, but are quite variable. |
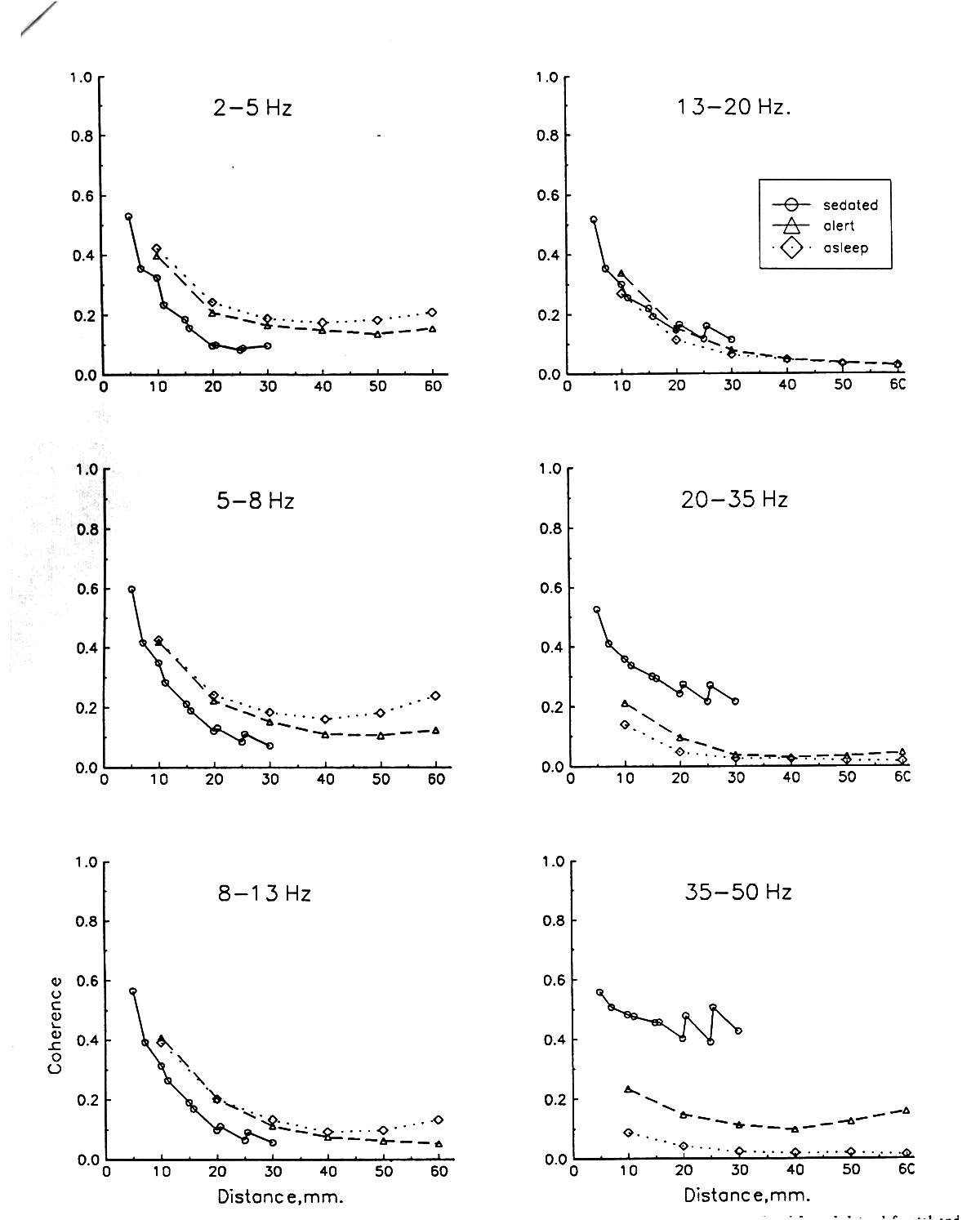 |
Figure 5. Coherence for several bands. Subdural recording, Patient K, subfrontal, lateral frontal and temporal cortex; as Fig. 4 but a more typical example of sleep, with very slight but consistently higher coherence in the lower bands. The sedated coherence is higher in the higher bands in this example, but not generally. Standard errors (not shown) are very small since the data are large. |
C. Coherence of adjacent pairs in a row
In order to investigate the basic question whether there is non-random structure at the resolution offered by available electrode spacing, we looked at the coherence values for adjacent pairs along each strip of subdural electrodes and along the depth electrode arrays. Graphs of the coherence for individual pairs along a strip of eight (Figs. 7, 8, 9) reveal local discontinuities rather than a smooth gradation of neighboring values - that is, the spatial frequency is wideband, including millimeter wave lengths. Successive pairs, for example made up of electrodes 1 & 2, 2 & 3, 3 & 4, up to 7 & 8 are separated by 10 mm in the usual subdural strips, 5 mm in the several cases with finer spacing. Note that adjacent pairs share one electrode.
 |
Figure 6. Coherence as function of distance, comparing pre-ictal and seizure states. Medial temporal depth electrode array; Patient W. Means of only 3-6 computing epochs of 10 s each. Most commonly all bands increase in coherence during a seizure; here only the three highest bands show a substantial increase. The 35-50 Hz band shows the standard errors; they are mostly smaller than the symbols. |
Two principal findings are these. (i) Coherence can change abruptly, within 5-10 mm, even when the pairs share a member. (ii) The profiles of coherence along a strip tend to be similar for different frequency bands.
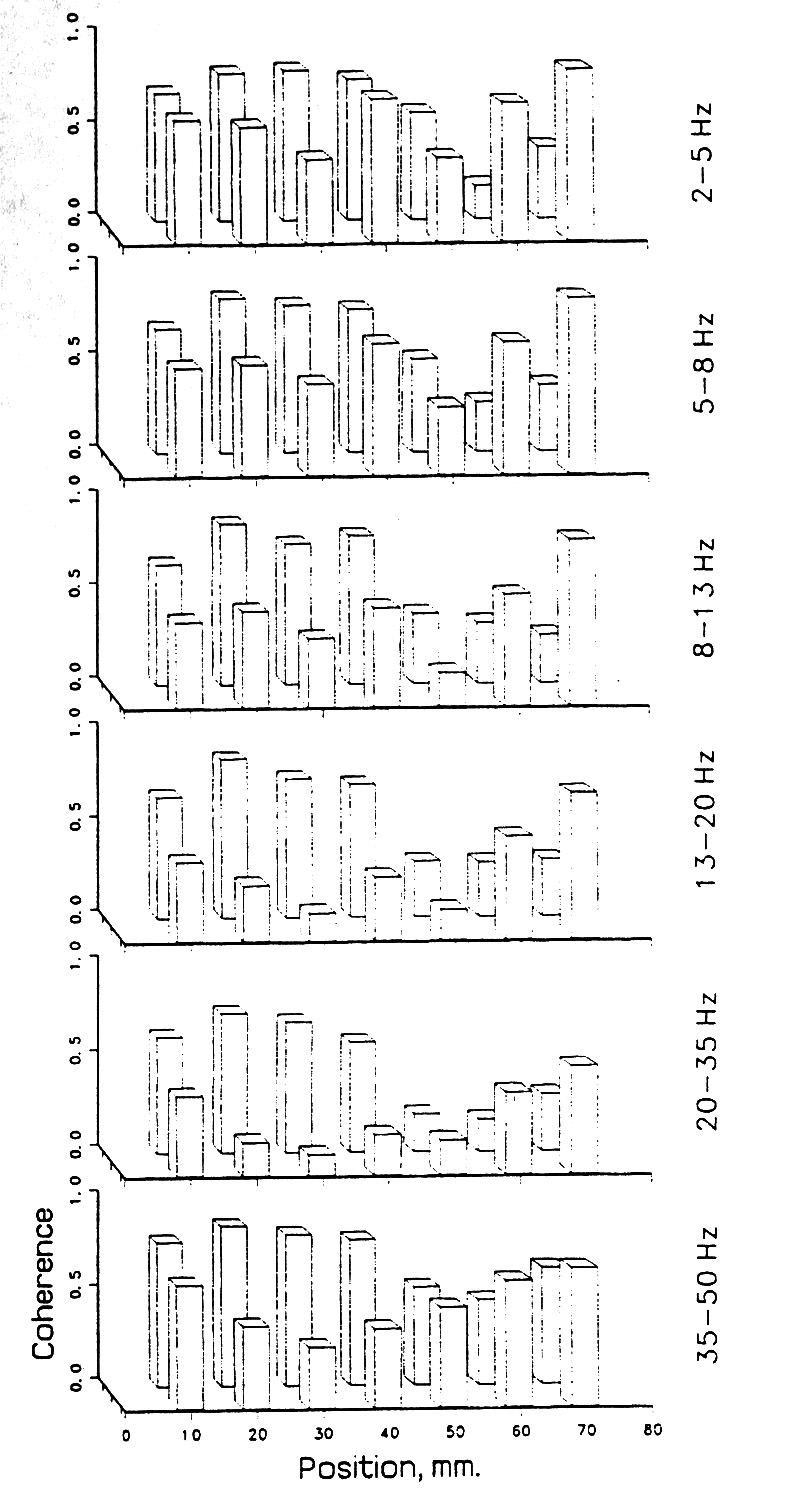 |
Figure 7. Local coherence; bar graph of the values for neighboring pairs, thus electrodes 1-2, 2-3, 3-4, ...7-8. Each bar is the mean of 18 epochs of 5 s each; subdural array on the lateral inferior occipital and posterior temporal cortex; Patient A. There were two rows of 8 electrodes with 10 mm spacing. The tendency is typical for a similar profile of the bars in successive bands. |
To evaluate the significance of the first of these findings, we consider two possible null hypotheses - both denying that there are significant structural features.
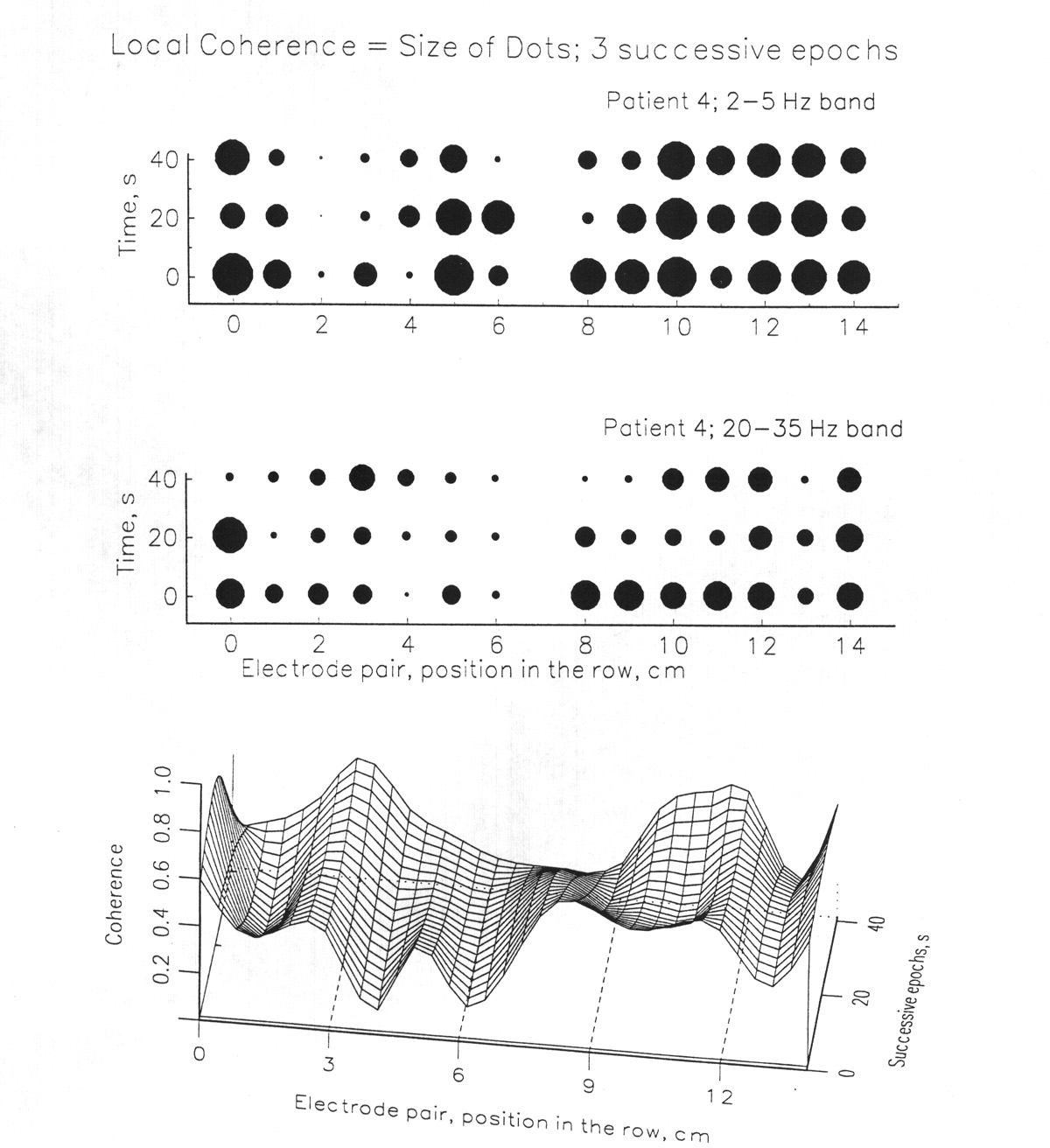 |
Figure 8. Local coherence; above, dot display of data similar to that in Fig. 7, arranged to show the abrupt discontinuities in space and time. The three rows of dots are successive 20 s epochs; subdural 10 mm grid array on parieto-occipital cortex in Patient Me. See text for statistical assessment. The two rows of electrodes are plotted end to end, not in their natural position. Below, a spline surface plot of the same data, which emphasizes the local features of coherence. Actually, the assumption of the interpolating algorithm is not justified and, instead, there may be even finer-grained ridges and valleys, both in the spatial (X) and the time (Y) dimensions. |
(i) Absence of fine structure would be expected if adjacent values are similar and, in the extreme case, form a simple gradient, e.g. from the highest to the lowest. This would be true if coherence tends to extend over such large domains, with low spatial frequency, (as one gathers from the literature on scalp coherence) that one cannot speak of local or fine structure. Our data rule out this alternative. Abrupt steps, cliffs, valleys and buttes are common, whereas the expectation from this hypothesis is the maximal number of minimal steps.
 |
Figure 9. Local coherence of neighboring pairs in the 2-5 Hz band, along a medial temporal depth array in Patient H. The four rows are successive epochs of only 3 s. Note that abrupt discontinuities in the spatial and temporal dimensions, as seen in subdural recordings extend into depth recordings of the hippocampus and adjoining structures. |
To quantify the discontinuities and compare the data with the null hypothesis of distribution of the coherence (C) values, we first computed the ratio (R) of the lower divided by the higher of each pair of adjacent (nearest neighbor) coherence values. Each strip of 8 electrodes gives 7 adjacent C values and 6 nearest neighbor pairs of C values for computing R values, which represent the size of a step or cliff in the profile, whether up or down. The N of such R values from the 6 subdural patients is 898. An R=0.8 signifies that the C between one pair of adjacent electrodes along a strip is 8/10ths of the C of the neighboring pair that shares one electrode; smaller Rs, like 0.1, are larger cliffs. These numbers, being ratios, are independent of the absolute level of coherence but when C is small, a small difference in C can be a large step in ratio (small R). The findings are based on all the suitable files of data from six subdural and 3 depth-recorded patients, each represented by one to two minutes of EEG in 20 s coherence estimates. Large steps (Rs of 0.5 and smaller) occur in the original data well in excess of their incidence in the null hypothesis of smooth, low spatial frequency C distribution, emulated by sorting the observed C values in order (ascending or descending) before calculating the nearest neighbor ratios. Small steps (Rs above 0.8) are much less common in the original data than in the ordered emulation of the low spatial frequency hypothesis. These relations between the incidence of R values in the data and the sorted series are quite consistent, although not invariable, among bands and patients. Figure 10 shows the incidence of R values in 10 classes for one patient in two bands, and the distribution on this and the other null hypothesis.
 |
Figure 10. Nearest neighbor ratios to test the incidence of abrupt steps in coherence between adjacent pairs. Data from subdural electrode strips on lateral frontal, subfrontal and inferior temporal cortex of Patient 6; two of the six bands are shown. "Data" means the observed distribution of values; "Sorted" means the original coherence values are arranged in order, e.g. from largest to smallest before computing the ratios of neighboring values; "Shuffled" means the order of the values, was rearranged by chance 1000 times. The first two are unique and have no variance; the 95% confidence intervals of the shufflings shows they most often differ from the data insignificantly in the single size classes but the small excess of one above and the other below ca. 0.5 is repeated in many bands and patients. |
(ii) The alternative null hypothesis of no significant structure is that the fluctuations in observed values are distributed at random. We shuffled the order of the observed coherence values within each strip 1000 times and then computed the ratios of adjacent pairs of values and the means and standard deviations of their incidence in each of the 10 classes of ratios, from 0.1 to 1.0, to compare with the original, unshuffled incidence. There are significant differences between the shuffled and the unshuffled only in a few of the size classes but the small excess of the Rs >0.5 in the unshuffled data and of Rs <0.5 in the shuffled series is present in all 6 patients compared, with numbers of neighboring pairs of coherence values varying from 60-209, including all bands and both awake and asleep samples. Figure 10 shows the incidence in one patient and two bands. The hypothesis of random distribution of coherence values can be rejected; there is a higher tendency than that for adjacent values to be similar. Significant local structure is a departure from both randomness and extremely low spatial frequency. The observed distribution is quite unlike both extreme cases - smoother than the stochastic model and more structured than the large-scale-without-features model.
We have not yet found an electrode spacing small enough that every local peak or valley of coherence has several adjacent, similar coherence values. It is not clear whether there is a maximum spatial frequency or a volume spread of EEG so that all pairs within a certain radius will have similar coherence to the common, inactive reference and hence high coherence with each other. Coherence of a given pair of electrodes 10 mm apart is poorly predicted by knowing the coherence of neighboring pairs, even those sharing one member, but it weakly correlates with them. So far we have not discerned any pattern in the incidence of discontinuities but the limitation to strips of 8 electrodes would make it hard to notice any pattern except one of a spatial frequency of 3-4 cm per cycle, oriented at right angles to the strip.
D. Coherence compared between regions, lobes and hemispheres
In most patients the subdural strips with a 7 cm row of 8 electrodes had been placed over two regions, such as the subfrontal and the lateral frontal cortex or the anterior and posterior temporal cortex, usually on both left and right sides. Sometimes as many as three major lobes were sampled, parietal, temporal and frontal; the occipital was rarely sampled. We looked for differences between regions, lobes and hemispheres by comparing the means of all possible pairs of the electrodes within each region or the means of the 10 mm pairs, which are the most abundant and have the highest coherence.
Some subjects showed no difference at all between the rows of electrodes over different regions, excluding seizure states (Patients #1, #6, #7, except for small differences in a few bands where the temporal lobe is slightly higher than lateral frontal in bands 3 & 4 but not different from subfrontal; ...). Others showed markedly higher coherence over one region or lobe (#2 anterior temporal >> lateral temporal in all bands, pooling sleep and waking states; #5 lateral frontal (l.f.) > inferior temporal (i.t.) in band 2, the reverse in band 1 in sample R01; l.f.>i.t. in bands 2-4, i.t.>l.f. in bands 1 & 6 in sample R02; l.f.>i.t. in bands 1-5 in sample R05, all awake ) Some showed no significant difference between hemispheres, comparing the approximately symmetrical placements of the strips (Patient #6; ...); others showed markedly higher coherence over one hemisphere, for all frequency bands, e.g. Patient #1 on the left; #2 on the left, pooling states, but not by a significant amount in the 3 lower bands, significant in bands 4 and 5, actually reversed in band 6 where right is significantly > left,; #5 right >> left, both in sleep and waking states; #7 left >> right in all bands in both waking and sleep states, except band 6, awake, which is insignificantly greater on the left. We did not find any regional differences that were consistent through the available sample of subjects.
Data from the patients with depth arrayss in the medial temporal lobe include some between lobes (hippocampus to ipsilateral inferior or middle temporal gyrus ) and across the midline (contralateral homotopic loci and contralateral heterotopic loci). Values are generally substantially lower than for the largest distances between loci along the array, most often below 0.1. The highest values are usually in the 5-8 Hz band and a modest upturn is not uncommon in the band above 35 Hz.
E. Coherence as function of state (sleeping, waking, seizure and sedated states)
To compare coherence between states, values computed epochs of one second were averaged over 20 epochs and these means were pooled with those for 3-6 consecutive 20 second periods for all pairs of electrodes into one value for each of six frequency bands, for a one or two minute EEG sample for a given patient, already read and categorized by the neurologist. Sleep or drowsiness, in stages 2-3, shows significantly higher coherence than the awake state in 18 out of 72 pairwise comparisons of the two states in the same subject (Table 3: 12 pairs of states times 6 frequency
| F band, Hz | 1: 2-5 | 2: 5-8 | 3: 8-13 | 4: 13-20 | 5: 20-35 | 6: 35-50 |
| Pat. #1 Sleep CS1 | 0.44 0.02 | 0.47 0.02 | 0.48 0.02 | 0.46 0.02 | 0.41 0.02 | 0.35 0.03 |
| " Sleep CS2 | 0.53 0.02 | 0.54 0.02 | 0.55 0.02 | 0.52 0.02 | 0.41 0.02 | 0.35 0.03 |
| " Awake CS3 | 0.44 0.02 | 0.47 0.03 | 0.44 0.02 | 0.46 0.02 | 0.39 0.02 | 0.34 0.03 |
| Pat. #2 Drowsy C02 | 0.40 0.04 | 0.46 0.05 | 0.46 0.05 | 0.44 0.05 | 0.31 0.05 | 0.52 0.05 |
| " Awake C01 | 0.65 0.04 | 0.64 0.04 | 0.65 0.04 | 0.60 0.04 | 0.39 0.04 | 0.54 0.04 |
| " Awake C03 | 0.72 0.03 | 0.74 0.03 | 0.70 0.03 | 0.66 0.03 | 0.45 0.04 | 0.59 0.03 |
| Pat. #3 Drowsy A1a | 0.36 0.03 | 0.31 0.03 | 0.21 0.03 | 0.15 0.03 | 0.12 0.03 | 0.34 0.03 |
| " Awake A1b | 0.23 0.04 | 0.23 0.03 | 0.19 0.03 | 0.14 0.03 | 0.13 0.03 | 0.39 0.02 |
|
Pat. #4 Drowsy M8 " M9 Awake M4 " M7 |
0.25 0.02 0.27 0.02 0.27 0.02 0.34 0.02 |
0.24 0.02 0.21 0.01 0.18 0.01 0.25 0.02 |
0.25 0.01 0.20 0.01 0.15 0.01 0.25 0.01 |
0.17 0.01 0.14 0.01 0.15 0.01 0.18 0.01 |
0.12 0.01 0.21 0.01 0.12 0.01 0.17 0.01 |
0.11 0.01 0.40 0.02 0.12 0.01 0.18 0.01 |
| Pat. #5 Sleep Not6 | 0.17 0.01 | 0.15 0.01 | 0.14 0.01 | 0.14 0.01 | 0.07 0.01 | 0.06 0.01 |
| " Awake All | 0.19 0.01 | 0.17 0.01 | 0.16 0.01 | 0.16 0.01 | 0.17 0.01 | 0.35 0.02 |
| Pat. #6 Sleep M4,5 | 0.15 0.01 | 0.16 0.01 | 0.15 0.01 | 0.08 0.01 | 0.05 <0.005 | 0.04 <0.005 |
| " Awake M3,6 | 0.12 0.01 | 0.13 0.01 | 0.11 0.01 | 0.07 0.01 | 0.06 0.01 | 0.04 <0.005 |
| Pat. #7 Sleep Ksl | 0.30 0.03 | 0.30 0.03 | 0.26 0.03 | 0.18 0.02 | 0.08 0.01 | 0.05 0.01 |
| " Awake Kaw | 0.34 0.03 | 0.31 0.03 | 0.32 0.03 | 0.23 0.03 | 0.15 0.02 | 0.17 0.02 |
bands). Sleep is significantly lower in coherence in 24 of the 72. The other 30 paired comparisons are not significantly different but most of them are lower in sleep. These comparisons are weakened by the fact that when we have two samples of the same state, taken some time apart, 14 out of 24 paired comparisons (12 for sleep and 12 for awake) are significantly different. Coherence of all six bands is found to be a poor correlate of the state, which was defined both by observing the patient and by the EEG as read by the neurologist. If we look only at the lower two bands, of the 24 paired states in the same patient, sleep is significantly higher in 8, lower in 7 and not different in 9; even the lower frequencies do not consistently show more coherence in sleep.
In some large files in one patient we compared the states in a different way. The difference in mean coherence for each band between sleeping and waking states for 2 min samples of EEG for each pair of electrodes was taken and the mean differences tested for significance by the paired samples T-test. This procedure also showed that some sleep samples are significantly different from some waking samples whereas others are not, and that some sleep samples are significantly different from others. Differences between bands also do not appear to fall into a pattern.
The turn-up in coherence at high frequency, remarked above, occurred in 3 out of 9 sleep records in 3 out of 7 patients, compared to 4 out of the same number of awake records, in 3 patients, two of them the same ones who showed turn-ups in sleep. This feature is also not characteristic of either state. One cannot tell by inspection of the EEG whether coherence is high or low - therefore whether synchrony is high or low, in the sense of a population or volume congruence in phase. Sleep, thought to represent more synchrony than wakefulness, at least in lower frequencies, can have higher or lower coherence but more often has lower coherence - in all bands.
Several files of data come from the operating room and are included because electrode grid arrays were used with the 5 mm spacing. Although the patients were under light systemic sedation, they were able to answer questions and obey commands. The EEG is characterized by Dr. V.J. Iragui as having high beta and considerable theta; delta can be low or high. The mean coherences for all pairs and some selected distances are given in Table 4. Evidently, this kind of sedation has little if any effect on the coherence at this scale.
| F bands: Hz | 1: 2-5 | 2: 5-8 | 3: 8-13 | 4: 13-20 | 5: 20-35 | 6: 35-50 |
| Pat. #6 All | 0.29 0.04 | 0.27 0.04 | 0.28 0.04 | 0.21 0.03 | 0.14 0.03 | 0.07 0.02 |
| Pat. #7 All | 0.26 0.03 | 0.31 0.03 | 0.29 0.03 | 0.30 0.02 | 0.45 0.03 | 0.64 0.02 |
| " 5 mm | 0.51 0.06 | 0.58 0.05 | 0.56 0.05 | 0.52 0.05 | 0.59 0.05 | 0.69 0.05 |
| " 10 mm | 0.29 0.05 | 0.32 0.05 | 0.29 0.04 | 0.30 0.05 | 0.44 0.07 | 0.63 0.07 |
| " 20 mm | 0.05 0.02 | 0.08 0.02 | 0.070.02 | 0.12 0.05 | 0.31 0.11 | 0.53 0.12 |
| Pat. #8 All | 0.18 0.01 | 0.14 0.01 | 0.21 0.02 | 0.25 0.02 | 0.17 0.01 | 0.06 0.01 |
|
" 5 mm " 10 mm |
0.38 0.04 0.18 0.03 |
0.36 0.04 0.13 0.02 |
0.52 0.04 0.22 0.04 |
0.60 0.04 0.26 0.04 |
0.44 0.02 0.18 0.01 |
0.17 0.02 0.06 0.01 |
The depth recordings from medial temporal loci show that coherence averaged over 3 seizure episodes, for 30 seconds or more each, almost always increases above that in the pre-seizure period by 0.05 to 0.2 in all bands except the 5-8 Hz band. Other occasional exceptions are no increase or even a decrease in the 8-13 Hz band and even in the 35-50 Hz band. Shorter samples of EEG during seizures vary widely, as expected from the profound changes both in form and localization of the activity between the moment which the electroencephalographer designates as the start of a seizure and the maximal electrical paroxysm. In the last named stage, when the maximum number of channels are involved, the coherence is close to maximal (but rarely 1.0) in the higher frequency bands - especially above 15-20 Hz.
|
|
Discussion |
|
|
|---|
General conclusions and interpretations of the findings
The usual explicit or implicit model of the compound field potentials of the brain provides only large scale congruence. This model is not supported by the facts that we commonly see coherence on average tending to decline with distance between the two loci - by half in the first 8-10 mm, and that local differentiation of single pairs, or discontinuities in rows of pairs, is common in this measure of cooperativity. Such observations represent dynamic structure in the millimeter domain. Of course, these findings do not conflict with the known facts that coherence is often well above the chance level over many centimeters in scalp recordings, especially in limited bands and states, such as high alpha, delta and seizure states. The findings emphasize that scalp recording blurs the spatial structure of cooperativity and that subdural recording as well as depth recording with macroelectrodes reveals local structure and discontinuities in coherence in the domain of a few millimeters.
The significance of the finding that coherence tends to vary from pair to pair of loci in parallel for all frequency bands between 2-5 Hz and 35-50 Hz is, first, that it conflicts with the common model of the EEG as a mixture of rhythms from independent oscillators. We wonder whether that model has been influenced by the convenience of the FFT method of analysis, which assumes such independence. Of course, rhythms do occur. But the clearly established oscillations are only five or six and each tends to occur under special states of the brain, usually accounting for only a fraction of the whole energy of the wideband EEG. Most of the EEG most of the time is not obviously rhythmic. The second significance of the coherence evidence is the suggestion that a major part of the ongoing activity seen by local macroelectrodes on the pial surface and in the depths in sleep and wakefulness, in the absence of large alpha, comes from wideband events such as steps, corners, triangles and nonsinusoidal transients, not strongly rhythmic recurrences.
The finding that about half of the EEG samples from this collection of subjects show a steadily declining coherence with higher frequency bands and half show a turn-up in the bands 20-35 and 35-50 Hz, usually quite sharply and steeply in the highest band suggests an episodic incidence of correlated (phase-locked) activity within the gamma band. Since coherence only tends to be correlated with power but does not necessarily covary with it, there need not be a new peak in the power spectrum. We are reminded of new findings by J.C. Prechtl (1994) in turtle cortex where gamma spindles and peaks of coherence at that frequency occur with shifts of gaze or changes of accommodation. Whatever causes these up-turns, it is not peculiar to the waking or the sleeping state; it resembles the same, nearly universal feature of seizure states and may be due to seizure features in the EEG not visible by inspection. The turn-ups occur, not only when no visible electrical seizure activity is in progress but in lobes of the brain remote from the seizure focus. It would be most desirable to have comparable recordings from nonepilepsy subjects, to look for the incidence of a rise in coherence in the gamma band.
Comparison with literature on human coherence
An extensive literature is available on EEG coherence in human subjects. To cite a limited selection, reports show local and hemisphere differences (Pockberger et al. 1985; Tucker et al. 1986; Nielsen et al. 1990; Boldyreva and Zhavoronkova 1991; Wang et al. 1992), sex differences (Marosi et al. 1993), changes with cognitive tasks (Tucker et al. 1985; Gasser et al. 1987; Rappelsberger and Petsche 1988), and changes in certain diseases (Morrison-Stewart et al. 1991; Schmid et al. 1992; Le Roc'h et al. 1993; Besthorn et al. 1994). These studies are generally done with scalp recording. Only epilepsy has been extensively studied with intracranial, subdural and intracerebral depth electrodes (citations in Introduction), but not with respect to the distribution of coherence. The general picture inferred from a vast amount of data is that coherence is a large scale property, expressing the synchrony of the EEG over centimeters and tens of centimeters. Additional literature, going back for decades, shows coherence between rostral and caudal poles of the cerebrum and between diencephalon and cortex. Such data did not exclude local features or discontinuities, of course, but it did not suggest fine structure like that demonstrated here.
Even finer structure now appears probable. Our earlier study on rats and rabbits (Bullock and McClune 1989) showed evidence that with intracortical extracellular microelectrodes and spacing in the range of tenths of a millimeter, one encounters abrupt changes between high and low coherence, perhaps even more often than pial surface or intracerebral macroelectrodes.
Comparisons with nonhuman species
Coherence has been little studied in laboratory animals. A few studies on rabbits concern other problems. Our own study (Bullock and McClune 1989 and later, unpublished observations) showed that samples of sleeping and waking EEG in rabbits and rats, if categorized by EEG inspection, have less variability than the present samples from epilepsy patients. Sleep rather consistently shows higher coherence in all bands in rabbits, but the difference is usually slight. We concluded that the apparent synchrony is generally due to the increase in power at low frequencies; it need not mean any increase in synchrony at all. Recruitment of generators or increased excursion of voltage in each generator, without any necessary change in synchrony, could and apparently does account for most of the observed increase in amplitude.
Our earlier study gave preliminary evidence that reptiles have somewhat less coherence at the same distance than laboratory mammals and that teleosts and elasmobranch fishes have still lower coherence. A single invertebrate, the gastropod Aplysia, in a number of sample recordings, was found to have no significant coherence between microelectrodes less than a millimeter apart in the neuropile, when loci dominated by single units were avoided. This apparent phylogenetic trend requires confirmation.
Limitations, caveats and future directions
The considerable inconsistency of coherence between subjects and even samples from the same subject, in both sleeping and waking EEGs might mean that these states were abnormal in these patients or in our samples. However, the experienced neurologist who read the EEGs did not note peculiarities beyond the normal range of variation among such patients. A more systematic study is needed before generalizing on differences between states in respect to the local structure.
The small sample of subjects and placements of electrodes and especially of states and instances of each state limits the representativeness of this study. It is sufficient for the conclusions we have permitted ourselves, some of which are rather strong or surprising. However, it needs to be extended both in numbers and in anatomical and functional regions examined as well as in the range
of brain or mental states.
Particularly needed are systematic studies of coherence and other signs and measures of cooperativity with intracerebral, extracellular microelectrodes. Limited samples of such recording, with fine tips only fractions of a millimeter apart have been reported by Elul (1967, 1968) and Bullock and McClune (1989), with results that are partly in agreement and partly not. Elul emphasized that such channels of ongoing micro-EEG are typically quite independent and noncongruous (he did not measure coherence but implied it is very low), even at small distances between tips. We repeated these experiments with fine glass pipettes in the rabbit and rat cortex and found low coherence values in some pairs but in most pairs coherence was quite to very high; the main impression was that variability in space and in time is higher than in surface recordings. Confirming Petsche et al. (1975, 1984) we noted marked differences in coherence with depth but did not find a consistent lamina of minimum coherence, as they reported for delta frequencies. The model we proposed was that surface electrodes are effectively macroelectrode recordings and integrate over a larger volume than fine electrodes in the parenchyma; the latter are able to see the local differences in activity in small groups of generators that really characterize laminae, nuclei, columns and neuropiles made of heterogeneous cells.
The time course of coherence is treated in another paper, in preparation, at the level of resolution of about one second, and is shown to have significant temporal structure, with nonrandom features that vary with place and state. According to preliminary experiments, other methods, such as nonlinear measures of higher moments - for example the bispectrum and bicoherence computation reveal additional nonrandom features of the spatial and temporal dynamic fine structure.
This work was supported by grants from the National Institutes of Neurological Diseases and Stroke
|
|
References |
|
|
|---|
Besthorn, C., Forstl, H., Geiger-Kabisch, C., Sattel, H., Gasser, T. and Schreiter-Gasser, U. EEG coherence in Alzheimer disease. Electroencephalogr. clin. Neurophysiol. 1994, 90: 242-245.
Boldyreva, G.N. and Zhavoronkova, L.A. Interhemispheric asymmetry of EEG coherence as a reflection of different functional states of the human brain. Biomed. Sci. 1991, 2: 266-270.
Bullock, T.H. and McClune, M.C. Lateral coherence of the electrocorticogram: a new measure of brain synchrony. Electroencephalogr. clin. Neurophysiol. 1989, 73:479-498.
Bullock, T.H., Iragui, V.J. and Alksne, J.F. Electrocorticogram coherence and correlation of amplitude modulation between electrodes both decline in millimeters in human as well as in rabbit brains. Soc. Neurosci. Abstr. 1990, 16: 1241
Bullock, T.H., Achimowicz, J.Z. and McClune, M.C. Coherence in human subdural EEG has fine structure in space and time. Soc. Neurosci. Abstr. 1992, 18: 906
Duckrow, R.B. and Spencer, S.S. Regional coherence and the transfer of ictal activity during seizure onset in the medial temporal lobe. Electroencephalogr. clin. Neurophysiol. 1992, 82: 415-422.
Elul, R. Statistical mechanisms in generation of the EEG. In: L.J. Fogel and F.W. George (Eds.), Progress in Biomedical Engineering, vol. 1. Spartan Books, Washington, D.C. 1967: 131-150.
Elul, R. Brain waves: intracellular recording and statistical analysis help clarify their physiological significance. In: Anonymous Data Acquisition and Processing in Biology and Medicine, Vol. 5 (Proceedings of the 1966 Rochester Conference). Pergamon Press, Oxford, 1968: 93-115.
Engel, J.J., Henry, T.R., Risinger, M.W., et al. Presurgical evaluation for partial epilepsy: relative contributions of chronic depth electrode recordings versus FDG-PET and scalp-sphenoidalictal EEG. Neurology, 1990, 40: 1670-1677.
Fein, G., Raz, J., Brown, F.F. and Merrin, E.L. Common reference coherence data are confounded by power and phase effects. Electroencephalogr. clin. Neurophysiol. 1988, 69: 581-584.
Garcia de Leon, M. Electrical source analysis by brain mapping techniques. Physiol. Measurement, 1993, 14 (Suppl.) 4A: A95-A98.
Gasser, T., Jennen-Steinmetz, J. and Verleger, R. EEG coherence at rest and during a visual task in two groups of children. Electroencephalogr. clin. Neurophysiol. 1987, 67: 151-158.
Kanner, A.M., Morris, H.H., Luders, H., Dinner, D.S., Van Ness, P. and Wyllie, E. Usefulness of unilateral interictal sharp waves of temporal lobe origin in prolonged video-EEG monitoring studies. Epilepsia, 1993, 34: 884-889.
Le Roc'h, K., Rancurel, G., Poitrenaud, J., Bourgin, P. and Sebban, C. Verbal fluency and EEG coherence in Alzheimer's disease. Neurophysiol. Clin. 1993, 23: 422-433.
Mackenzie, R.A., Matheson, J.M., Smith, J.S. and Dwyer, M. Surgery for refractory epilepsy. Med. J. Aust. 1990, 153: 69-72, 76.
Marosi, E., Harmony, T., Becker, J., et al. Sex differences in EEG coherence in normal children. Int. J. Neurosci., 1993, 72: 115-121.
Mihara, T., Matsuda, K., Tottori, T., et al. Conditions for omitting invasive long-term monitoring before surgical resection in patients with temporal lobe epilepsy. Jpn. J. Psychiatry Neurol., 1992, 46: 323-329.
Morrison-Stewart, S.L., Williamson, P.C., Corning, W.C., Kutcher, S.P. and Merskey, H. Coherence on electroencephalography and aberrant functional organisation of the brain in schizophrenic patients during activation tasks. Br. J. Psychiatry, 1991, 159: 636-644.
Nielsen, T., Abel, A., Lorrain, D. and Montplaisir, J. Interhemispheric EEG coherence during sleep and wakefulness in left- and right-handed subjects. Brain & Cognition, 1990, 14:113-125.
Pockberger, H., Petsche, H., Rappelsberger, P., Zidek, B. and Zapotoczky, H.G. On-going EEG in depression: a topographic spectral analytical pilot study. Electroencephalogr. Clin. Neurophysiol. 1985, 61: 349-358.
Prechtl, J.C. Visual motion induces synchronous oscillation in turtle visual cortex. Submitted
Rappelsberger, P. The reference problem and mapping of coherence: a simulation study. Brain Topography, 1989, 2: 63-72.
Rappelsberger, P. and Petsche, H. Probability mapping: power and coherence analyses of cognitive processes. Brain Topogr. 1988, 1: 46-54.
Schmid, R.G., Tirsch, W.S., Rappelsberger, P., Weinmann, H.M. and Poppl, S.J. Comparative coherence studies in healthy volunteers and Down's syndrome patients from childhood to adult age. Electroencephalogr. clin. Neurophysiol. 1992, 83: 112-123.
Tucker, D.M., Dawson, S.L., Roth, D.L. and Penland, J.G. Regional changes in EEG power and coherence during cognition: intensive study of two individuals. Behav. Neurosci. 1985, 99: 564-577.
Tucker, D.M., Roth, D.L. and Bair, T.B. Functional connections among cortical regions: topography of EEG coherence. Electroencephalogr. clin. Neurophysiol. 1986, 63: 242-250.
Wang, G., Takigawa, M. and Matsushita, T. Correlation of alpha activity between the frontal and occipital cortex. Jpn. J. Physiol. 1992, 42: 1-13.