FOOTNOTES
(1) Brief relative to the lifespan of the animal; for most primates, less than one year. return
(2) "...any investment by the parent in an individual offspring that increases the offspring's chance of surviving (and hence reproductive success) at the cost of the parent's ability to invest in other offspring." (Trivers 1972, p. 139).return
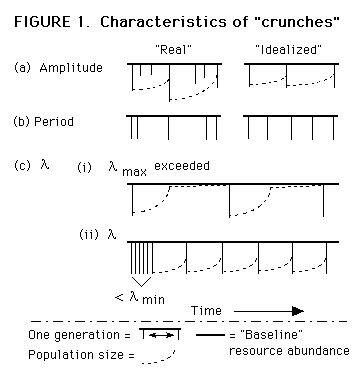
3 If V(L) is great relative to L and L is longer than a few years, it may become impossible to "plan" for minima by either method. As V(L) increases, however, the biologically relevant parameter will eventually become a function of the average interval between clusters of minima.
return
4 Many atoll peoples also make very explicit attempts to control their own fertility and are fully conscious of K (Bayliss-Smith 1974); conscious and unconscious strategies are expected to coexist in humans.
return
REFERENCES
Barkow, J. H., Burley, N. (1980). Human fertility, evolutionary biology, and the demographic transition. Ethol. Sociobiol. 1, 163-180.
Bayliss-Smith, T. (1974). Constraints on population growth: the case of the Polynesian outlier atolls in the pre-contact period. Hum. Ecol. 2, 259-295.
Birdsell, J. B. (1953). Some environmental and cultural factors influencing the structuring of Australian Aboriginal populations. Am. Nat. 87, 171-207.
Birdsell, J. B. (1975). A preliminary report on new research on man-land relations in Aboriginal Australia. In (A. C. Swedlund, Ed.) Population Studies in Archaeology and Biological Anthropology: A Symposium. Am. Antiq. 40: pt. 2 memoir 30. pp. 34-37.
Blurton Jones, N., Sibly, R. M. (1978). Testing adaptiveness of culturally determined behaviour: Do Bushmen women maximize their reproductive success by spacing births widely and foraging seldom ? In (V. Reynolds and N. Blurton Jones, Eds.), Human Behaviour and Adaptation. Symposium No. 18 of the Society for the Study of Human Biology. London: Taylor and Francis, pp. 135-157.
Broom, L., Siegel, B. J., Vogt, E. Z., Watson, J. B. (1967). Acculturation: an exploratory formulation. In (P. Bohannon and F. Plog, Eds.), Beyond the Frontier. New York: Natural History Press, pp. 255-286.
Cant, J. G. H. (1980). What limits primates ? Primates 21, 538-544.
Cohen, M. N. (1977). The Food Crisis in Prehistory. New Haven: Yale University Press.
Dolhinow, P., McKenna, J. J., vonder Haar Laws, J. (1979). Rank and reproduction among female langur monkeys: Aging and improvement (They're not just getting older, they're getting better). Aggressive Behavior 5, 19-30.
Draper, P. (1976). Social and economic constraints on child life among the !Kung. In (R. B. Lee and I. DeVore, Eds.) Kalahari Hunter-gatherers. Cambridge: Harvard University Press, pp. 199-217.
Durham, W. H. (1976). The adaptive significance of cultural behavior. Hum. Ecol. 4, 89-201.
Gaulin, S. J. C., Konner, M. (1977). On the natural diet of primates, including humans. In (R. J. Wurtman and J. J. Wurtman, Eds.), Nutrition and the Brain, Vol. 1. New York: Raven Press, pp. 1-86.
Hallowell, A. I. (1949). The size of Algonkian hunting territories: a function of ecological adjustment. Am. Anthropol. 51, 35-45.
Hamilton, W. D. (1964). The genetical evolution of social behaviour I, II. J. theor. Biol. 7, 1-52.
Hardesty, D. L. (1977). Ecological Anthropology. New York: Wiley.
Hartung, J. (1976). On natural selection and the inheritance of wealth. Current Anthropology 17, 607-622.
Hayden, B. (1972). Population control among hunter/gatherers. World Arch. 4, 205-221.
Hayden, B. (1975). The carrying capacity dilemma: an alternative approach. In (A. C. Swedlund, Ed.) Population Studies in Archaeology and Biological Anthropology: A Symposium. Am. Antiq. 40: pt. 2 memoir 30. pp. 11-21.
Howell, N. (1979). Demography of the Dobe !Kung. New York: Academic Press.
Just, P. (1980). Time and leisure in the elaboration of culture. J. Anthropol. Res. 36: 105-115.
Lee, R. B. (1968). What hunters do for a living, or how to make out on scarce resources. In (R. B. Lee and I. DeVore, Eds.) Man the Hunter. Chicago: Aldine, pp. 30-48.
Lee, R. B. (1980). Lactation, ovulation, infanticide, and women's work: a study of hunter-gatherer population regulation. In (M. N. Cohen, R. S. Malpass and H. G. Klein, Eds.) Biosocial Mechanisms of Population Regulation. New Haven: Yale University Press, pp. 321-348.
Radcliffe-Brown, A. R. (1948). The Andaman Islanders. Glencoe: The Free Press.
Ricklefs, R. E. (1973). Ecology. Portland: Chiron Press.
Ripley, S. (1980). Infanticide in langurs and man: adaptive advantage or social pathology ? In (M. N. Cohen, R. S. Malpass and H. G. Klein, Eds.) Biosocial Mechanisms of Population Regulation. New Haven: Yale University Press, pp. 349-390.
Riss, D., Goodall, J. (1977). The recent rise to the alpha-rank in a population of free-living chimpanzees. Folia Primatol. 27, 134-151.
Schoener, T. W. (1971). Theory of feeding strategies. Ann. Rev. Ecol. Syst. 2, 369-404.
Slobodkin, L. B., Rapoport, A. (1974). An optimal strategy of evolution. Q. Rev. Biol. 49, 181-200.
Smith, E. A. (1979). Human adaptation and energetic efficiency. Hum. Ecol. 7, 53-74.
Southwood, T. R. E. (1977). Habitat, the templet for ecological strategies ? J. Anim. Ecol. 46, 337-365.
Stearns, S. C. (1976). Life-history tactics: a review of the ideas. Q. Rev. Biol. 51, 3-47.
Trivers, R. L. (1972). Parental investment and sexual selection. In (B. G. Campbell, Ed.) Sexual Selection and the Descent of Man 1871-1971. Chicago: Aldine, pp. 136-179.
Vayda, A. P., McCay, B. J. (1975). New directions in ecology and ecological anthropology. Ann. Rev. Anthropol. 4, 293-306.
Werner, O., Flowers, N. M., Ritter, M. L., Gross, D. R. (1979). Subsistence productivity and hunting effort in native South America. Hum. Ecol. 7, 303-315.
Wiens, J. A. (1977). On competition and variable environments. Am. Scientist 65, 590-597.
Wilmsen, E. N. (1979). Diet and fertility among Kalahari Bushmen. Working Papers No. 14 of the African Studies Center, Boston University. pp. 1-23.
Wilson, D. J. (1981). Of maize and men: a critique of the maritime hypothesis of state origins on the coast of Peru. Am. Anthropol. 83, 93-120.
Winterhalder, B. (1980). Environmental analysis in human evolution and adaptation research. Hum. Ecol. 8, 135-170.