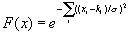
Published in Psycoloquy 9(67), 1998.
This paper presents a speculative model of the cognitive mechanisms underlying the transition from episodic to mimetic (or memetic) culture with the arrival of Homo erectus, which Donald [1991] claims paved the way for the unique features of human culture. The model draws on Kauffman’s [1993] theory of how an information-evolving system emerges through the formation of an autocatalytic network. Though originally formulated to explain the origin of life, this theory also provides a plausible account of how discrete episodic memories become woven into an internal model of the world, or worldview, that both structures, and is structured by, self-triggered streams of thought. Social interaction plays a role in (and may be critical to) this process. Implications for cognitive development are explored.
abstraction, animal cognition, autocatalysis, cognitive development, cognitive origins, consciousness, cultural evolution, memory, meme, mimetic culture, representational redescription, worldview.
1. The subject of cultural origins is usually approached from an archeological perspective. For example, by dating artifacts such as tools we learn approximately when humans acquired the ability to make and use those tools. This paper takes a more cognitive approach [see also Barkow et al. 1992; Donald 1991, 1993a, 1993b; Tomasello et al. 1993; Tooby & Cosmides 1989]. It outlines a theory of the psychological mechanisms underlying the major cognitive transition that, Donald [1991] proposes, made possible the characteristic complexity and ingenuity of human culture.
2. The theory proposed here was inspired by an idea originally put forward to explain the origin of life. The origin of life and the origin of culture might appear at first glance to be very different problems. However at a gross level of analysis they amount to the same thing: the bootstrapping of a system by which information patterns self-replicate, and the selective proliferation of some variants of these self-replicating patterns over others. The theory is thus consistent with the perspective of culture as a form of evolution [Dawkins 1975; Gabora 1997]. In keeping with this evolutionary framework, the term ‘meme’ is used to refer to a unit of cultural information as it is represented in the brain. Thus ‘meme’ refers to anything from an idea for a recipe to a memory of one’s uncle to a concept of size to an attitude of racial prejudice. The rationale for lumping together episodic memories and symbolic abstractions is that they are both ‘food for thought’, units of information that can be drawn upon to invent new memes or to clarify existing ones. Memes that have been implemented as actions, vocalizations, or objects are referred to as artifacts.
3. The basic line of reasoning in this paper goes as follows. The bottleneck to cultural evolution appears to be the capacity for a self-sustained stream of thought that both structures and is structured by an internal model of the world, or worldview. It is this capacity that enables us to plan and predict, to generate novelty, and to tailor behavior according to context. The problem is: until discrete memories have been woven into a conceptual web, how can they generate a stream of thought? And conversely, until a mind can generate a stream of thought, how does it weave its memories into a worldview? Kauffman’s proposal that life originated with the self-organization of a set of autocatalytic polymers suggests a mechanism for how this comes about. Much as catalysis increases the number of different polymers, which in turn increases the frequency of catalysis, reminding events increase meme density by triggering symbolic abstraction, which in turn increases the frequency of remindings. And just as catalytic polymers undergo a phase transition to a state where there is a catalytic pathway to each polymer present, and together they constitute a self-replicating set, memes undergo a phase transition to a state where each meme is retrievable through a pathway of remindings/associations, and together they constitute a transmittable worldview. In the origin of life scenario, since reactions occur in parallel, autocatalytic closure increases sharply as the ratio of reactions to polymers increases. In the cultural analog, however, the retrieval and invention of memes is funneled through an attention/awareness mechanism, which introduces a bottleneck. Therefore, this transition occurs gradually, as increasingly abstract concepts are perceived and their implications percolate through the memetic network. Social interaction and artifacts facilitate the process, and ensure that the continued evolution of memes does not hinge on the survival of any particular meme host.
4. This section will present background material relevant to the central thesis. We begin with a comparison of minds that are and are not able to sustain cultural evolution, and draw on what is known about human cognition to make some hypotheses concerning the differences between them. We then turn to the paradox of the origin of life, and show how autocatalysis provides a potential solution.
5. In Origins of the Modern Mind, Donald [1991] argues convincingly that the capacity for abstract thought is the bottleneck of cultural evolution, and that it came about during the transition from episodic to mimetic culture following the arrival of Homo erectus approximately 1.7 million years ago. Before this time the human memory system was like that of a primate [1], limited to the storage and cued retrieval of specific episodes. Accordingly Donald uses the term episodic to designate a mind in which episodic memory is the only memory system there is. An episodic mind is capable of social attribution, insight and deception, and is sensitive to the significance of events. With much training it can learn arbitrary stimulus-response associations (such as pointing at a token of a certain shape to obtain food). However it can not invent symbols or abstractions on its own, or experiment with them. It has great difficulty accessing memories independent of environmental cues, and is unable to improve skills through self-cued rehearsal.
6. In contrast, the mimetic mind has, built upon its episodic foundations, a "multimodal modeling system with a self-triggered rehearsal loop". In other words, it can retrieve and recursively operate on memories independent of environmental cues, a process referred to by Karmiloff-Smith [1992] as representational redescription. By redescribing an episode in terms of what is already known, it gets rooted in the network of understandings that comprise the worldview, and the worldview is perpetually restructured as new experiences are assimilated, and new symbols and abstract concepts invented as needed. A mimetic individual is able to rehearse and refine skills, and therefore exhibits enhanced behavioral flexibility, and more precise control over intentional communication. The upshot is cultural novelty. "Mime, play, games, toolmaking, and reproductive memory," says Donald "are thus manifestations of the same superordinate mimetic controller." The appearance of sophisticated stone tools, long-distance hunting strategies, and migration out of Africa, as well as the rapid increase in brain size at this time [Bickerton 1990; Corballis 1991; Lieberman 1991], are cited as evidence for the transition from episodic to mimetic culture. Donald claims "it is not clear that the mimetic controller must be localized in any single anatomical structure, although it must have functional unity". Mimetic ability seems to encompass a broad panoply of skills associated with several distinct regions of the brain. Since miming accounts for only a small part of what the mimetic mind can do, and since mimetic skill seems to boil down to the capacity to evolve memes, we will use the term memetic instead of mimetic.
7. Donald’s proposal is invaluable in that spurs us to consider the cognitive basis of culture, but it leaves us hanging as to what sort of functional reorganization could turn an episodic mind into a memetic one. In particular, it leaves us with a nontrivial problem of origins. In the absence of representational redescription, how are relationships established amongst memes so that they become a worldview? And until a memory incorporates relationships between stored items, how can one meme evoke another which evokes another, et cetera, in a stream of representational redescription? We know that the brains of an ancestral tribe somehow turned into instruments for the variation, selection, and replication of memes. What happened to get the ball rolling, to enable the process of memetic evolution to take hold? When Groga, a member of this tribe, had her first experience, there were no previously-stored episodes to be reminded of, just external and internal stimuli (such as hunger). As episodes accumulated in her memory, occasionally it happened that an instant of experience was so similar to some stored episode that a retrieval process occurred, and she was reminded of that past episode. Perhaps the retrieval elicited a learned response. For example, the sight of a bumpy, red gourd might have reminded her of a bumpy, yellow gourd that her brother once used to carry water. Embedded in this recollection was the refreshing taste of the water he shared with her. The memory might have inspired her to use the red gourd to carry water. But since her memory consisted only of stored episodes, no abstractions, this is the only kind of influence it could exert; her awareness was dominated by the stimuli of the present moment. At some point in her life, however, she managed to willfully direct her attention not to a particular sensory stimulus, nor to the performance of a biological drive satisfying action, but to a chain of symbol manipulation. She kept this stream of thought going long enough to refine a concept or perspective, or invent a novel artifact. But if you need an interconnected worldview to generate a stream of thought, and streams of thought are necessary to connect individual memes into a worldview, how could one have come into existence without the other?
8. The first step toward an answer is to elucidate as best we can given present knowledge, the cognitive mechanisms that distinguish a memetic mind from an episodic one. This section describes a minimal, biologically-plausible cognitive architecture that could qualify as memetic. This cognitive architecture is a ‘best guess’, drawn from evidence in cognitive science, neuroscience, and artificial intelligence and the knowledge that the memetic mind grew out of (and therefore the potential for it was implicit in) the architecture of the episodic mind. It has the following attributes. First, it can integrate inputs from the sensorium and the drives with inputs from memory, and dispense commands to the motor system. Second, the memory is sparse, content addressable, distributed, modular, and habituates to repeated inputs. This enables it to generate abstractions. Finally, it can manipulate abstractions, consciously and recursively. We discuss each attribute briefly, explain how together they accomplish a memetic task, and then specify what is most likely lacking in the episodic mind.
9. Integration of memory, stimuli, and drives. Vital to both the episodic and the memetic mind is a means of integrating sensations, drives, and stored memories to produce a seamless stream of conscious experience and purposeful motor action. The place where this information is coordinated need not correspond to a single anatomical structure (though it is often suggested that the intralaminar nuclei of the thalamus are involved). We will adopt Kanerva’s [1988] term, the focus, since it does not imply any commitment regarding centrality or global penetration. We assume that the states of the neurons that comprise the focus determine the content and phenomenal qualities of an instant of awareness. A meme is then a high-dimensional vector of difference relations (or continuous variables) that either is or has been encoded in an individual’s focus.
10. Sparse. Our sensory apparatus can register a tremendous amount of information. Where n is the number of features the senses can distinguish, N, the number of memes that could potentially be hosted by the focus = 2n for boolean variables (and it is infinitely large for continuous variables). For example, if n =1,000, N = 21,000 memes [2]. Assuming n is large, N is enormous, so the memory is sparse in that the number of locations L where memes can be stored is only a small fraction of the N perceivable memes. In other words, neural pathways leading out from the focus do not receive inputs from each of its n slots, but from some fraction of them. The number of different memes actually stored at a given time, s, is constrained by L. The set of all possible n-dimensional memes a mind is capable of storing can be represented as the set of vertices (if features assume only binary values) or points (if features assume continuous values) in an n-dimensional hypercube, where the s stored memes occupy some subset of these points. The distance between two points in this space is a measure of how dissimilar they are, referred to as the Hamming distance. Kanerva [1988] makes some astute observations about this memory space. The number of memes at Hamming distance d away from any given meme is equal to the binomial coefficient of n and d, which is well approximated by a Gaussian distribution. Thus, if meme X is 111...1 and its antipode is 000...0, and we consider meme X and its antipode to be the ‘poles’ of the hypersphere, then approximately 68% of the other memes lie within one standard deviation (sqrt[n]) of the ‘equator’ region between these two extremes (FIGURE 1). As we move through Hamming space away from the equator toward either Meme X or its antipode, the probability of encountering a meme falls off sharply by the proportion sqrt[n]/n.
FIGURE 1. Solid black curve is a schematic distribution of the Hamming distances from address of a given meme to addresses of other memory locations in a sparse memory. The Gaussian distribution arises because there are many more ways of sharing an intermediate number of features than there are of being extremely similar or different. A computer memory stores each item in only the left-most address, whereas a distributed network stores it throughout the network. A restricted activation function, such as the radial basis function, is intermediate between these two extremes. Activation decreases with distance from the ideal address, as indicated by green shading.
11. In fact the space of possibilities is even larger if we assume that the mind rarely if ever attends all the stimulus dimensions it is capable of detecting. Therefore the number of dimensions the focus pays attention to, n, is only a subset of the maximum, M; i.e. n £ M. The strength of the signal on a neural pathway from memory, senses, or drives must surpass some threshold before the dimension of the focus it activates is attended. Since the memory can now store memes of any length up to M, the number of possible memes is:
| N = 2M+1 - 2 » 2M+1 | (1) |
The bottom line is: the memory would probably have to be larger than the number of particles in the universe to store all the permutations of sensory stimuli it is capable of registering. It is therefore sparse.
12. Distributed Representation. In a sparse memory, the probability that a given meme in the focus is identical to one in storage is virtually zero, which means that retrieval should be impossible. In connectionist networks, this problem is solved by distributing the storage of a meme across many locations. Likewise, each location participates in the storage of many memes. The focus is represented as input/output nodes, memory locations as hidden nodes, and their pattern of connectivity as weighted links. An input touches off a pattern of activation which spreads through the network until it relaxes into a stable configuration, or achieves the desired input-output mapping using a learning algorithm. The output vector is determined through linear summation of weighted inputs. Thus a retrieved meme is not activated from a dormant state, but ‘reconstructed’. This approach is necessary if we aim to model cognition at a fine-grained level of resolution—down to the threshold of human discrimination. There is a saying ‘You never step into the same stream twice’, and this saying applies to streams of thought as well as streams of water. Right now I am retrieving a memory of eating cinnamon toast, and tomorrow I may retrieve the ‘same’ memory. But today it is colored by today’s mood, today’s events; tomorrow it will experienced slightly differently. It isn’t the exact same information pattern conjured up time and again. The reconstructive approach enables the memory to abstract a prototype, fill in missing features of a noisy or incomplete pattern, or create a new meme on the fly that is more appropriate to the situation than any meme it has actually experienced [Rumelhart & McClelland 1986]. For example, if an autoassociative network has been fed vectors in which feature one is present whenever feature two is present, and vice versa, it will respond to an input that lacks information about feature one, such as *101, by generating 1101. It may never have actually encountered 1101 before, but given that in its ‘world’ there exists a correlation between features one and two, this is an appropriate response. In addition to associations between inputs and outputs of features, the network has learned a higher-level association between two features. In effect, it contains more information than has been fed into it.
13. A problem with distributed representation is: unless stored patterns are perfectly orthogonal, they interfere with one another, a phenomenon known as crosstalk. This is solved by restricting the storage region. For instance, in Kanerva’s [1988] Sparse Distributed Memory (SDM) model, a meme is stored in all locations within a hypersphere of addresses surrounding the ideal address. The smaller the Hamming distance between two memes, the more their storage locations overlap, so the higher the probability they are retrieved simultaneously and blended in the focus. A more sophisticated way of implementing this idea, for which there is neurobiological evidence, is to use a radial basis function (RBF) [Clothiaux et al 1991; Hancock et al 1991; Willshaw & Dayan 1990]. Once again a hypersphere of locations is activated, but this time activation is maximal at the center of the RBF and tapers off in all directions according to a (usually) Gaussian distribution (see FIGURE 1). Where x is an i-dimensional input vector, kis the center of the RBF, and s is the width of the Gaussian, hidden nodes are activated as follows:
|
|
(2) |
By carving out a hypersphere in memory space, one part of the network can be modified without affecting the capacity of other parts to store other patterns. The further a stored meme is from k, the less activation it not only receives but in turn contributes to the next evoked meme, and the more likely its contribution is cancelled out by that of other memes. In neural networks, suitable values for k and s are found during a training phase. In the brain, k values could be modified by changing the pattern of neuron interconnectivity. Decreasing neuron activation thresholds would increase s.
14. Organized Modularity. Another way of avoiding cross-talk is to induce a division of labor amongst competing subnetworks; in other words, to make the memory modular [Nowlan 1990; Jacobs et al. 1991]. There is abundant evidence of modularity in the brain, and its preservation in phylogenetic history suggests that it is not arbitrary. We assume that (1) the world we live in is highly patterned and redundant, and (2) this pattern and redundancy is reflected in the connectivity of the neurons where memes are stored. After birth there is a large-scale pruning of neurons. It seems reasonable that the surviving subset of the M possible inputs to each neural pathway is determined by biological and cultural selective pressures, instead of at random. These pressures sculpt the pattern of neuron connectivity such that the L (out of N possible) locations can store most of the memes we stand a chance of encountering. This means that in practice the sparseness of the memory does not interfere with its representational capacity. It also means that the probability a given stimulus activates a retrieval event is not as low as the statistics suggest.
15. Content addressable. A computer reads from memory by simply looking at the address in the address register and retrieving the item at the location specified by that address. The sparseness of human memory prohibits this kind of one-to-one correspondence. However, content addressability can be feigned, as follows. The feature pattern that constitutes a given meme causes some neurons leading out from the focus to be excited, and others inhibited. The ensuing chain reaction activates memory neurons where the meme gets stored. The address of a memory neuron amounts to the pattern of excitatory and inhibitory synapses from focus to storage that make it fire, so there is a systematic relationship between the information content of a meme and the locations it activates. Thus, embedded in the neural environment that supports their informational integrity, memes act as implicit pointers to other memory locations. These pointers prompt the dynamic reconstruction of the next meme to be subjectively experienced, which is statistically similar to the one that prompted it. As a result, the entire memory does not have to be searched in order for a gourd to remind Groga of a previously-encountered gourd. It is worth stressing that there is no search taking place, just information flowing through a system displaced from equilibrium. The current instant of experience activates certain neurons, which in turn activate certain other neurons, which leads to the distributed storage of that experience, which activates whatever else is stored in those locations, which then merges with any salient information from the senses and drives to form the next instant of experience, et cetera in an ongoing cycle. What emerges is that the system appears to retrieve memories that are similar, or concepts that are relevant, to the current experience. But that’s not magic; it is simply a side effect of the fact that correlated memes get stored in overlapping locations.
16. Habituation. We do not want an ongoing stimulus, such as the sound of rain, to recursively evoke remindings of rain. The nervous system avoids this kind of perseveration as follows. First, neurons have a refractory period during which they can not fire, or their response is greatly attenuated. Second, they ‘team play’; the responsibility for producing a response is shared by a cooperative group of neurons such that when one is refractory another is active. If exactly the same neurons are stimulated repeatedly, they all become refractory, and there is little or no response.
17. Capacity for Symbol manipulation. The connectionist methods described above are examples of the subsymbolic approach to cognition, which works best for modeling perceptual and low-level cognitive phenomena. These include detecting, representing, and responding flexibly to patterns of correlation, learning fuzzy categories, and solving simple constraint satisfaction problems. Subsymbolic processing makes the world easier to navigate. But the world contains additional structure that our brains are not hardwired to capture. Therefore, even after memes are stored in memory, they are clustered, rather than being uniformly distributed throughout the space of possible memes, and contain implicit predicate logic relationships. This is where symbolic processing is useful. Symbolic models of cognition focus on the serial and potentially recursive application of logical operations on symbols, without attempting to represent their internal structure. It is particularly good at modeling the high-level cognitive abilities that are unique to memetic minds, such as planning and deductive reasoning. Arguments for a reconstructive view of retrieval notwithstanding, highly abstract concepts that have been used thousands of times, such as ‘space’ or ‘equal’ or ‘is’, would be unlikely to emerge from memory retaining the associations of any particular usage. Thus it seems reasonable to begin with the working hypothesis that subsymbolic processing predominates for low-level, parallel, automatically-generated cognitive phenomena, and that symbolic processing provides a satisfactory approximation for many high-level, serial, consciously-directed aspects of cognition. (Creative processes may draw heavily on both.)
18. Let us now examine how a cognitive architecture with these attributes would accomplish a specific task. Consider the situation wherein the sight of a wrotting, striped, bumpy, red gourd reminds Groga of the striped, bumpy, yellow gourd her brother used as to carry water, which generates the desire to have water readily available in the cave. Groga slashes the top off the red gourd and scoops water into it. To her dismay, the water leaks out through a soft decay spot. Just out of sight lies the intestine of a recently-killed water buffalo. What sort of cognitive dynamics would prompt Groga to tie one end of the intestine and use it as a waterbag? It is unlikely that the ability to classify ‘gourd’ and ‘knotted intestine’ as potentially-substitutable instances of the category ‘container’ is hardwired. No one in Groga’s tribe has previously conceived of an intestine as a container, so social learning is not an option. This task involves a number of difficult skills including abstract reasoning, uncued retrieval, redescription, and manual dexterity. It lies beyond the horizon of what the episodic mind can accomplish.
19. The sight of the red gourd is registered as a vector of features in her focus. This vector determines which synapses leading out from the focus are excited and which are inhibited, which determines how activation flows through her memory network, which in turn determines the hypersphere of locations where ‘red gourd’ is stored. The process of storing to these neurons triggers retrieval from these neurons of whatever has been stored in them. Of course, nothing is retrieved from them if, after ‘red gourd’ is stored, Groga’s attention is directed toward some stimulus or biological drive. But to the extent that memory contributes to the next instant of awareness, storage of ‘red gourd’ activates retrieval of not only ‘red gourd’ itself but all other memes stored in the same locations. The next meme to be encoded in the focus is found by evaluating the contributions of all retrieved memes feature-by-feature. Whereas the retrieved copies of ‘red gourd’ reinforce one another, the other retrieved memes contribute less, and are statistically likely to cancel one another out. They do not cancel out exactly, however, unless the distribution of stored memes within the hypersphere of activated locations is uniformly dense. In this case it is not. The meme ‘yellow gourd container’, which got stored when she saw her brother carrying water in a yellow gourd, acts as an attractor. The result is that the next meme ends up being ‘red gourd container’. Though it is a reconstructed blend, something Groga has never actually experienced, it can still be said to have been retrieved from memory.
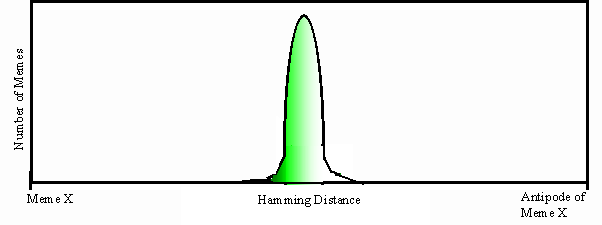
20. Groga pours water in the red gourd and, as we know, it leaks out. Her mental model of the world was in error; not all gourds can transport water. Stymied, memory is probed again, with knowledge of relationships between objects and attributes guiding the process. The second probing occurs with intensified activation of the pathway leading from the ‘concave’ slot of the focus, and inhibition of the ‘permeable’ slot. Let us now zero in on the portion of Groga’s memory that deals with four discrete features—bumpy, striped, permeable, and concave (FIGURE 2). ‘Bumpy’, ‘striped’, ‘permeable’ and ‘concave’ lie on the x1, x2, x3, and x4 axes respectively, and a black dot represents the center of a distributed hypersphere where a meme is stored. The second probing of memory activates a slightly different set of locations, which evoke the abstract category ‘container’, the class of objects that are concave and impermeable, and for which the attributes bumpy and striped are irrelevant. ‘Container’ was implicit in the meme-space; it covered the two-dimensional yellow region of the original hypercube. More generally, we can view an n-dimensional meme space as a set of nested hypercubes, such that implicit in the outermost hypercube of memes with all n dimensions there exist hypercubes of memes with n-1 dimensions, n-2 dimensions, et cetera. Armed with the category ‘container’, Groga dips into memory again to discover what else constitutes a member of this category. The closest thing she can come up with is ‘intestine’. Symbol manipulation now kicks in. She realizes that the intestine is impermeable and almost concave. Knotted at one end it, ‘intestine’ would constitute another member of the category ‘container’. She could therefore carry water in it. She runs off to fetch the intestine.
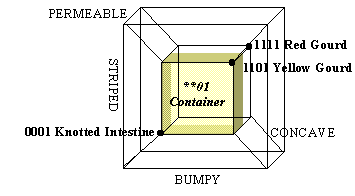
Figure 2. A four-dimensional hypercube representing a segment of memory space. ‘Bumpy’, ‘striped’, ‘permeable’ and ‘concave’ lie on the x1, x2, x3, and x4 axes respectively. Three memes are stored in the space: ‘intestine’, ‘yellow gourd’, and ‘red gourd’. Black dots represent the centers of the distributed hyperspheres where they are stored. The category ‘container’ occupies the central yellow region. To make use of this implicit abstraction, it is necessary to recognize that ‘concave’ and ‘permeable’ are the relevant dimensions. An impermeable gourd can be used as a container by cutting off the top, and an intestine can be used as a container by tying a knot at one end.
21. The foregoing discussion may be wrong in the details, but hopefully it captures the gist of memetic cognition. Now we ask: what is the episodic mind lacking? Some are tempted to say that the ability of animals to respond appropriately to salient stimuli, and even learn arbitrary sensorimotor associations, indicates some capacity for symbolic thought. However animals’ learned behavior is stereotyped and brittle—it can not be adapted to new contexts—which suggests that they use symbols only in an iconic sense. They give no indication of engaging in streams of thought that reorganize memes in ways that make their similarities and differences more explicit. They could not retrieve the memory that an intestine is in the cave, much less realize it is relevant to the goal of transporting water. Our best-guess model of cognition suggests a number of possible reasons. First, the resolution of the perceptual apparatus might not be high enough to capture enough features of salient stimuli (large M). Second, there might not be enough memory locations to keep these distinctions intact during storage (large L). Third, the density s/N of stored memes might be too low. In other words, there might not be enough different basins of attraction for memes to slide into, or not enough of these attractors are occupied. Another possibility is that the neuron activation threshold is too high (and thus s too narrow). The end result is the same in all cases: rarely is there a stored meme within retrievable distance of a given meme in the focus. Thus the memory does not encode relationships, so rarely can a stream of interrelated thoughts ensue. In fact, these explanations are connected. M limits L, which in turn limits s. And since if s = 0, the memory only retrieves memes identical to the content of the focus and therefore can not form abstractions, s also limits s.
22. At this point we are in a position to reframe our central question. We want to know how a mind comes to assume a self-sustained stream of thought that progressively shapes and is shaped by, a worldview. Abstract thinking requires each meme that enters the focus to activate one or more memes already stored in memory enough to evoke a retrieval. The memory must be traversed with tunnels that connect related concepts like an apple crisscrossed with wormholes. However, representational redescription is the process that puts related memes within working memory reach of one another; it is what recognizes abstract similarities and restructures the memory to take them into account. How do you get the wormholes without the worms?
23. We will put the aside question of cultural origins for now, and turn to the problem of biological origins. The paradox of the origin of life can be stated simply: if living things come into existence when other living things give birth to them, how did the first living thing arise? That is, how did something complex enough to reproduce itself come to be? In biology, self-replication is orchestrated through an intricate network of interactions between DNA, RNA, and proteins. DNA is the genetic code; it contains instructions for how to construct various proteins. Proteins, in turn, both catalyze reactions that orchestrate the decoding of DNA by RNA, and are used to construct a body to house and protect all this self-replication machinery. Once again, we have a chicken-and-egg problem. If proteins are made by decoding DNA, and DNA requires the catalytic action of proteins to be decoded, which came first? How could a system composed of complex, mutually dependent parts come into existence?
24. The most straightforward explanation is that life originated in a ‘prebiotic soup’ where, with enough time, the right molecules collided into one another at the same time and reacted in exactly the right ways to create the DNA-RNA-protein amalgam that is the crux of life as we know it. Proponents argue that the improbability of this happening does not invalidate the theory because it only had to happen once; as soon as there was one self-replicating molecule, the rest could be copied from this template. Miller [1955] increased the plausibility of this hypothesis by showing that amino acids, from which proteins are made, form spontaneously when a reducing [3] mixture of oxygen, hydrogen, carbon, nitrogen, water, and ammonia is subjected to high energy. These molecules were all likely to have been present on the primitive earth, and energy could have come in the form of electric discharges from thunderstorms, ultraviolet light, or high temperatures generated by volcanoes. Other experiments have shown that the molecular constituents of DNA and RNA, as well as the fatty acids from which membranes are constructed, can be formed the same way. Unfortunately, the complexity of the DNA-RNA-protein structure is so great, and in the earth’s early atmosphere the concentrations of the necessary molecules were so dilute, that the probability of life originating this way is infinitesimally low. Hoyle and Wickramasinghe [1981] likened it to the probability that a tornado sweeping through a junkyard would spontaneously assemble a Boeing 747.
25. The less complex something is, the more feasible its spontaneous generation. The discovery of ribozymes—RNA molecules that, like proteins, are capable of catalyzing chemical reactions—brought hope that first living molecule had been found. With ribozymes you wouldn’t need DNA or proteins to establish a self-replicating lineage; these RNA molecules would do the job of all three. In practice, however, self-replication of RNA is fraught with difficulties. It tends to fold back on itself creating an inert, tangled mess [Joyce, 1987]. Furthermore, the probability of a ribozyme assembling spontaneously from its components is remote [Orgel 1987], and even if it managed to come into existence, in the absence of certain error-detecting proteins found in all modern-day organisms, its self-replication capacity would inevitably break down in the face of accumulated error over successive generations [Eigen and Schuster, 1979]. Thus it is far from obvious how the chain of self-replicating systems that eventually evolved into you and I got started.
26. Despite the myriad difficulties encountered attempting to get ribozymes to self-replicate, the idea behind it—that life originated in a simple self-replicating system that over time evolved into the familiar DNA-RNA-protein complex—was a good one. Once you have some sort of self-replicating structure in place, anything whatsoever that accomplishes this basic feat, natural selection can enter the picture and help things along. Kauffman [1991] suggested that knowing as much as we do about what life is like now may actually get in the way of determining how it began. Accordingly, he decided to focus on how to get from no life at all to any kind of primitive self-replicating system, and hand the problem of getting from there to DNA-based life, over to natural selection (as well as self-organizing processes). Given the conditions present on earth at the time life began, how might some sort of self-replicating system have arisen? His answer is that life may have begun not with a single molecule capable of replicating itself, but with a set of collectively self-replicating molecules. That is, none of the molecules could replicate itself, but each molecule could induce the replication of some other molecule in the set, and likewise, its own replication was induced by some other member of the set. This kind of dual role as both ingredient (or stimulant) and product of different chemical reactions is not uncommon for polymers such as protein and RNA molecules.
27. Polymers induce each other’s replication by acting as catalysts. Catalysts speed up chemical reactions that would otherwise occur very slowly. An autocatalytic system is a set of molecules which, as a group, catalyze their own replication. Thus if A catalyzes the conversion of X to B, and B catalyzes the conversion of Y to A, then A + B comprise an autocatalytic set (FIGURE 3). In an environment rich in X and Y, A + B can self-replicate. A set of polymers wherein each molecule’s formation is catalyzed by some other molecule is said to exhibit catalytic closure.
FIGURE 3. An autocatalytic set: A catalyses the formation of B, and B catalyses the formation of A. Thick black arrows represent catalyzed reactions. Thin green arrows represent catalysis.
28. It is of course highly unlikely that two polymers A and B that just happened to bump into one another would happen to catalyze each other. However, this is more likely than the existence of a single polymer catalyzing its own replication. And in fact, when polymers interact their diversity increases, and so does the probability that some subset of the total reaches a critical point where there is a catalytic pathway to every member. To show that this is true we must show that the number of reactions by which they can interconvert increases faster than their total number. Given polymers made up of, say, two different kinds of monomers, of up to a maximum length of M monomers each, then N, the number of polymers, is 2M+1 as per equation (1). Thus as M increases—which it obviously does, since two of the longest polymers can always join to form a longer one—the number of polymers increases exponentially. Now we need to show that the number of reactions between them increases even faster. We will be conservative and consider only cleavage (e.g. 110 -> 1 + 10) and ligation (e.g. 1 + 10 -> 110) reactions on oriented polymers (such as protein and RNA fragments). The number of possible reactions R is the product of the number of polymers of a certain length times the number of bonds, summed across all possible lengths:
| R | = 2M(M - 1) + 2M-1(M - 2) + ... +2M-(M-2)(M - (M - 1)) | |
| (3) |
Dividing equation (3) by equation (1), we find that as M increases, the ratio of reactions to polymers increases by a factor of M-2. This means that if each reaction has some probability of getting carried out, the system eventually undergoes a transition to a state where there is a catalytic pathway to each polymer present. The probability of this happening shifts abruptly from highly unlikely to highly likely as R/N increases. This kind of sharp phase transition is a statistical property of random graphs and related systems such as this one. Random graphs consist of dots, or ‘nodes’, connected to each other by lines or ‘edges’. As the ratio of edges to nodes increases, the probability that any one node is part of a chain of connected nodes increases, and chains of connected nodes become longer. When this ratio reaches approximately 0.5, almost all these short segments become cross-connected to form one giant cluster (FIGURE 4). Plotting the size of the largest cluster versus the ratio of edges to nodes yields a sigmoidal curve. The larger the number of nodes, the steeper the vertical portion of this curve (referred to as the percolation threshold).
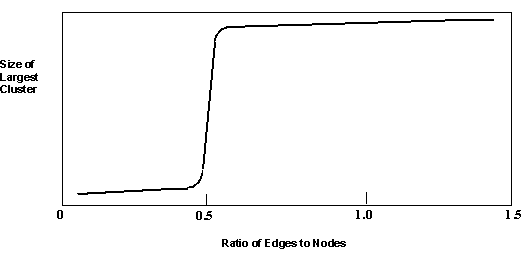
FIGURE 4. When the ratio of edges to nodes reaches approximately 0.5, short segments of connected nodes join to form a large cluster that encompasses the vast majority of nodes.
29. Of course, even if catalytic closure is theoretically possible, we are still a long way from knowing that it is the correct explanation for the origin of life. How likely is it that an autocatalytic set would have emerged given the particular concentrations of chemicals and atmospheric conditions present at the time life began? In particular, some subset of the R theoretically possible reactions may be physically impossible; how can we be sure that every step in the synthesis of each member of an autocatalytic set actually gets catalyzed? Kauffman’s response is: if we can show that autocatalytic sets emerge for a wide range of hypothetical chemistries—i.e., different collections of catalytic molecules, then the particular details of the chemistry that produced life do not matter so long as it falls within this range. We begin by noting that, much as several different keys sometimes open the same door, each reaction can be catalyzed by not a single catalyst, but a hypersphere of catalytic molecules, with varying degrees of efficiency. So we assign each polymer an extremely low a priori random probability P of catalyzing each reaction. The lower the value of P, the greater M must be, and vice versa. Kauffman shows that the values for M and P necessary to achieve catalytic closure with a probability of > 0.999 are highly plausible given the conditions of early earth. Experimental evidence for this theory using real chemistries [Lee et al. 1996, 1997; Severin et al. 1997], and computer simulations [Farmer, et al. 1986] have been unequivocally supportive. Farmer et al. showed that in an ‘artificial soup’ of information strings capable of cleavage and ligation reactions, autocatalytic sets do indeed arise for a wide range of values of M and P. FIGURE 5 shows an example of one of the simplest autocatalytic sets it produced. The original set of polymers from which an autocatalytic set emerges is referred to as the ‘food set’. In this case it consists of 0, 00, 1, and 11. As it happens, the autocatalytic set that eventually emerges contains all members of the original food set. This isn’t always the case.
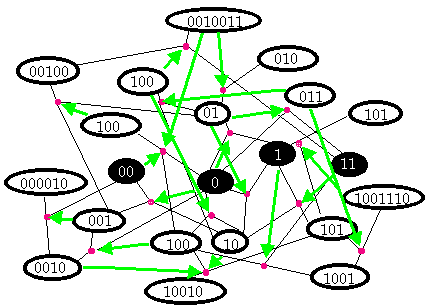
FIGURE 5. A typical example of a small autocatalytic set. Reactions are represented by thin black lines connecting ligated polymers to their cleavage products. Thick green lines indicate catalysis. Dark ovals represent food set.
30. An interesting question explored in this simulation is: once a set of polymers has achieved autocatalytic closure, does that set remain fixed, or is it able to incorporate new polymer species? They found that some sets were subcritical—unable to incorporate new polymers—and others were supracritical—incorporated new polymers with each round of replication. Which of these two regimes a particular set fell into depended on P, and the maximum length of the food set polymers.
31. Now the question is: supposing an autocatalytic set did emerge, how would it evolve? The answer is fairly straightforward. It is commonly believed that the primitive self-replicating system was enclosed in a small volume such as a coascervate or liposome to permit the necessary concentration of reactions [Oparin 1971; Morowitz 1992; Cemin & Smolin, in press]. Since each molecule is getting duplicated somewhere in the set, eventually multiple copies of all molecules exist. The abundance of new molecules exerts pressure on the vesicle walls. This often causes such vesicles to engage in a process called budding, where it pinches off and divides into two ‘twins’. So long as each twin contains at least one copy of each kind of molecule, the set can continue to self-replicate indefinitely. Replication is far from perfect, so an ‘offspring’ is unlikely to be identical to its ‘parent’. Different chance encounters of molecules, or differences in their relative concentrations, or the arrival of new ‘food’ molecules, could all result in different catalysts catalyzing a given reaction, which in turn alters the set of reactions to be catalyzed. So there is plenty of room for heritable variation. Error catastrophe is unlikely because, as mentioned earlier, initially each reaction can be catalyzed not by a single catalyst but by a hypersphere of potential catalysts, so an error in one reaction does not have much effect on the set at large [4]. Selective pressure is provided by the affordances and limitations of the environment. For example, say an autocatalytic set of RNA-like polymers arose. Some of its offspring might have a tendency to attach small molecules such as amino acids (the building blocks from which proteins are made) to their surfaces. Some of these attachments inhibit replication, and are selected against, while others favor it, and are selected for. We now have the beginnings of the kind of genotype-phenotype distinction seen in present-day life. That is, we have our first indication of a division of labor between the part of the organism concerned with replication (in this case the RNA) and the part that interacts with the environment (the proteins).
32. The autocatalysis origin of life theory circumvents the ‘chicken-and-egg’ problem by positing that the same collective entity is both code and decoder. This entity doesn’t look like a code in the traditional sense because it is a code not by design but by default. The code is embodied in the physical structures of the molecules; their shapes and charges endow them with propensities to react with or ‘mutually decode’ one another such that they manifest external structure, in this case a copy of its collective ‘self’. Since autocatalytic sets appear to be a predictable, emergent outcome in any sufficiently complex set of polymers, the theory suggests that life is an expected outcome rather than a lucky long-shot.
33. We have taken a look at two paradoxes—the origin of culture and the origin of life—which from hereon will be referred to as OOC and OOL respectively.The parallels between them are intriguing. In each case we have a self-replicating system composed of complex, mutually interdependent parts, and since it is not obvious how either part could have arisen without the other, it is an enigma how the system came to exist. In both cases, one of the two components is a storehouse of encoded information about a self in the context of an environment. In the OOL, DNA encodes instructions for the construction of a body that is likely to survive in an environment like that its ancestors survived. In the OOC, an internal model of world encodes information about the self, the environment, and the relationships between them. In both cases, decoding a segment of this information storehouse generates another class of information unit that coordinates how the storehouse itself gets decoded. Decoding DNA generates proteins that, in turn, orchestrate the decoding of DNA. Retrieving a memory or concept from the worldview and bringing it into awareness generates an instant of experience, a meme, which in turn determines which are the relevant portion(s) of the worldview to use in constructing the next instant of experience. In both cases it is useful to think of the relevant class of information units as states in an information space, each of which can act on a hypersphere of other states. In the memory model it was the hypersphere of related memes, and in the OOL model it was the hypersphere of potential catalysts.
34. We have argued that the most likely bottleneck in the OOC is the establishment of a network of inter-related memes, a worldview, that progressively shapes and is shaped by a stream of self-triggered thought. We want to determine how such a complex entity might come to be. Donald claims that the transition from episodic to memetic culture "would have required a fundamental change in the way the brain operates." Drawing from the OOL scenario presented above, we will explore the hypothesis that meme evolution begins with the emergence of a collective autocatalytic entity that acts as both code and decoder. This idea was mentioned briefly in [Gabora 1996a; 1996b; 1997]; here it is fleshed out it in greater detail.
35. In the OOL case we asked: what was lying around on the primitive earth with the potential to form some sort of self-replicating system? The most promising candidate was catalytic polymers, the molecular constituents of either protein or RNA. Here we ask an analogous question: what sort of information unit does the episodic mind have at its disposal? It has memes, specifically memories of episodes. Episodic memes then constitute the ‘food set’ of our system.
36. Next we ask: what happens to the ‘food set’ to turn it into a self-replicating system? In the OOL case, food set molecules catalyzed reactions on each other that increased their joint complexity, eventually transforming some subset of themselves into a collective web for which there existed a catalytic pathway to the formation of each member molecule. An analogous process might concievably transform an episodic mind into a memetic one. Food set memes activate redescriptions of each other that increase their joint complexity, eventually transforming some subset of themselves into a collective web for which there exists a retrieval pathway to the formation of each member meme. Much as polymer A brings polymer B into existence by catalyzing its formation, meme A brings meme B into conscious awareness by evoking it from memory. As in Section II.2, a ‘retrieval’ can be reminding, a redescription of something in light of new contextual information, or a creative blend or reconstruction of many stored memes.
37. How might Groga’s mind have differed from that of her ancestors such that she was able to initiate this kind of transformation? In the OOL case, it was crucial that the polymers be catalytic. We simply gave each polymer a small, random probability P of catalyzing each reaction. In the OOC case, we assume that each of Groga’s L memory locations where the s memes are stored has a RBF with a Gaussian distribution of width s centered on it. Thus the probability that one meme evokes another is determined s by rather than a random probability P. Let us consider what would happen if, due to some genetic mutation, Groga’s activation threshold were significantly lower than average for her tribe. Thus s is wider, which means that a greater diversity of memes are activated in response to a given experience, and a larger portion of the contents of memory merge and surface to awareness in the next instant. Since the memory is content-addressable, when meme X goes fishing in memory for meme X', sooner or later this large hypersphere is bound to ‘catch’ a stored meme that is quite unlike X. For example, let us say that Groga sees rabbits every day, so there are lots of rabbit memories stored in her brain. For simplicity, let us say they consist of a sequence of ten 0’s followed by a five bit long variable sequence. She happens to look off in the distance and see a grazing water buffalo, which gets represented in her focus as 000000011101010. The buffalo meme will be referred to as meme X. Because the hypersphere is wide, all of the rabbit memories lie close enough to meme X to get evoked in the construction of X' (as is X itself). Since all the components from which X' is made begin with a string of seven zeros, there is no question that X' also begins with a string of seven zeros. These positions might code for features such as ‘has eyes’, ‘eats’, et cetera. The following set of three 1s in the rabbit memes are canceled out by the 0s in the buffalo memes, so in X' they are represented as *s. These positions might code for features such as ‘floppy ears’. The last five bits constituting the variable region are also statistically likely to cancel one another out. These code for other aspects of the experience, such as, say, the color of the sky that day. So X' turns out to be the meme 0000000********, the generic category ‘animal’, which then gets stored in memory in the next iteration. This evocation of ‘animal’ by the buffalo episode isn’t much of a stream of thought, and it doesn’t bring her much closer to an interconnected conceptual web, but it is an important milestone. It is the first time she ever derived a new meme from other memes, her first creative act.
38. Although lowering the neuron activation threshold was what enabled Groga to create an abstraction, the penalty for having too low a threshold would be very high, because successive thoughts would not necessarily be meaningfully related to one another. Abstract thought, unlike episodic thought, can not rely on the continuity of the external world (i.e., if a desk is in front of you now it is likely to still be in front of you now) to lend coherence to conscious experience. Too low a threshold might be expected to result in a cognitive rendition of superconductivity, where lowering resistance increases correlation distance and thus a perturbation to any one pattern percolates through the system and affects even distantly-related patterns. (The free-association of a schizophrenic seems to correspond to what one might expect of a cognitive system with this property [see Weisberg 1986].) However, if the threshold is extremely high, such that distributions don’t overlap, the attended meme must be identical to one stored in memory to evoke a retrieval. To produce a steady stream of meaningfully-related yet potentially creative remindings, the threshold must fall within an intermediate range. This is consistent with Langton’s [1992] finding that the information-carrying capacity of a system is maximized when its interconnectedness falls within a narrow regime between order and chaos.
39. Thus thoughts do not leap from one unexplored territory of meme-space to another, but meander from one meme to a similar one in a region that has proven fruitful and is therefore exceptionally clustered with memes. This not only increases the frequency of remindings and abstractions, it provides a thread of continuity linking one meme to the next. Organized modularity also enhances continuity by precluding activation of irrelevant memes. Since statistical similarity is preserved across sequentially evoked memes in a train of thought, thinking can be viewed as an internal form of meme self-replication. It could be argued that the correlation between consecutive memes is so low that this hardly deserves to be called a form of self-replication. One wouldn’t want consecutive memes to be identical. Surely Eigen and Schuster’s error catastrophe argument applies here; that is, the ‘copying fidelity’ of this process is so low that errors would quickly accumulate and in no time the ‘lineage’ would ‘die’. But this argument doesn’t apply. The only reason it is a pitfall for biological evolution is that copying error tends to impair the capacity to self-replicate. So long as offspring are as good as their parents at reproducing themselves, and live long enough to do so, it doesn’t matter how much error is introduced from one generation to the next. It is only when a generation dies without having reproduced that is there a problem. In the biological world, once something is dead it can not spring forth life [5]. But in memetic evolution this isn’t necessarily the case. To show why this is so, say that half-way through the train of consecutive memes in Einstein’s brain that culminated in the theory of relativity, a tiger burst in through the window. The correlation between the relativity meme of one instant and the tiger-perception-meme of the next instant would be almost zero. This momentous memetic lineage would come to a screeching halt. But would it be lost forever? No. Sooner or later, once the tiger situation was taken care of, the relativity stream of thought would inevitably resume itself. Memory (and external artifacts) function as a memetic sperm bank, allowing a defunct ancestral line to be brought back to life and resume self-replication. The upshot is that in culture you can get away with a much higher error rate than in biology.
40. Cultural evolution has not only an internal form of replication, but also an internal means of generationg variation. In a stream of thought, consecutive memes aren’t exact replicas; each meme is a variation of its predecessor. It also has an internal form of selection. Selection comes in the form of drives, needs, attention-focusing mechanisms, and the associative organization of memory, which constrain how one meme evokes another. Thus all the components of an evolutionary process take place in the mind of an isolated individual. The memory-driven generation of a stream of correlated memes can be viewed as a coevolutionary relationship between replication, variation, and selection, and the process of representational redescription can itself be redescribed as the selective generation of variant replicants. Embedded in the outer, inter-individual sheath of memetic evolution we find a second intra-individual sheath, where the processes of replication, variation, and selection are not spatiotemporally separated but intimately intertwined. Together they weave a stream of thought, one meme fluidly transmuting into the next.
41. Thus the semantic continuity of a stream of thought makes memory navigable despite its sparseness. Once ‘animal’ has been evoked and stored in memory, the locations involved habituate and become refractory (so, for instance, ‘animal’ does not recursively evoke ‘animal’). However locations storing memes that have some animal features, but that were not involved in the storage of ‘animal’, are still active. Thus ‘animal’ might activate ‘tiger’ which might evoke ‘hienna’ et cetera, strengthening associations between the abstract category and its instances. Other abstractions, such as ‘container’, form in analogous fashion. As Groga accumulates both episodic memes and abstractions, the probability that any given attended meme is similar enough to some previously-stored meme to activate it increases. Therefore reminding acts increase in frequency, and eventually become streams of remindings, which get progressively longer. Groga is now capable of a train of thought. Her focus is no longer just a spot for coordinating stimuli with action; it is now a forum for abstractive operations that emerge through the dynamics of iterative retrieval.
42. We have seen how our best-guess model of human cognition achieves a stream of thought. How do we know that streams of thought will induce a phase transition to a critical state where for some subset of memes there exists a retrieval pathway to each meme in the subset? In the OOL case, we had to show that R, the number of reactions, increases faster than N, the number of polymers. We found that R/N increased by a factor of M–2, where M was the maximum number of monomers per polymer. Because of the highly parallel nature of this system, it was reasonable to equate potential reactions with actual reactions, and therefore to assume that the new polymers resulting from these reactions actually exist (and can themselves partake in reactions). Similarly, we now want to show that some subset of the memes stored in an individual’s mind inevitably reach a critical point where there is a path by which each meme in that subset can get evoked. But here, it is not reasonable to assume that all N perceivable memes actually exist (and can therefore partake in retrieval operations). Their number is severely curtailed by the number of memory locations, the variety of perceptual experience, and the fact that meme retrieval, though distributed at the storage end, is serial at the awareness end. The rate at which streams of thought reorganize the memetic network is limited by the fact that everything is funneled through the focus; we can only figure one thing out at a time. This presents a bottleneck that was not present in the OOL scenario. As a result, whereas OOL polymers underwent a sharp transition to a state of autocatalytic closure, any analogous transition in inter-meme relatedness is expected to take place gradually. So we need to show that R, the diversity of ways one meme can evoke another, increases faster than not N but s, the number of memes that have made it through this bottleneck. That is, as the memory assimilates memes, it comes to have more ways of generating memes than the number of memes that have explicitly been stored in it.
43. This brings us to another complication, which further prolongs cognitive development. Since short, simple molecules are more abundant and readily-formed than long, complex ones, in the OOL case it made sense to expect that the food set molecules were the shortest and simplest members of the autocatalytic set that eventually formed. Accordingly, in simulations of this process the ‘direction’ of novelty generation is outward, joining less complex molecules to form more complex ones through AND operations (see FIGURE 5). In contrast, the memetic food set molecules are complex, consisting of all attended features of an episode. In order for them to form an interconnected web, their interactions tend to move in the opposite direction, starting with relatively complex memes and forming simpler, more abstract ones through OR operations. The net effect of the two is the same: a network emerges, and joint complexity increases. But what this means for the OOC is that there are numerous levels of autocatalytic closure, which convey varying degrees of worldview interconnectedness and consistency on their ‘meme host’. These levels correspond to increased penetration of the (n-1, n-2…)-dimensional nested hypercubes implicit in the memory space. Since it is difficult to visualize a set of nested, multidimensional hypercubes, we will represent this structure as a set of concentric circles, such the outer skin of this onion-like structure represents the hypercube with all n dimensions, and deeper circles represent lower-dimensional hypercubes (FIGURE 6). Obviously, not all the nested hypercubes can be shown. The points of our original hypercube are represented as points along the perimeter of these circles, and k values (centermost location where a meme is stored) are shown as large, black dots. The outermost shell encodes memes in whatever form they are in the first time they are consciously encountered. It is all the episodic mind has to work with. In order for one meme in this shell to evoke another, they have to be extremely similar at a superficial level. In a memetic mind, however, related concepts are within reach of one another because they are stored in overlapping hyperspheres.
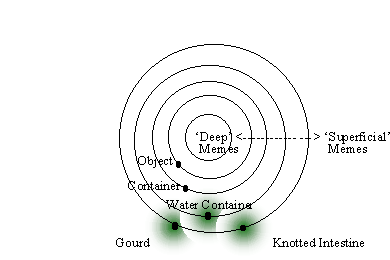
FIGURE 6. The role of abstractions in creative thought. For ease of visualization, the set of nested hypercubes representing the space of possible memes is shown as a set of concentric circles, where deeper circles store deeper layers of abstraction (lower dimensional hypercubes). A black dot represents the centermost storage location for a specific meme. ‘Water container’ is a more general concept than ‘gourd’ or ‘knotted intestine’, and is therefore stored at a deeper layer. Green circle around each stored meme represents hypersphere where the meme gets stored and from which the next meme is retrieved. ‘Gourd’ and ‘knotted intestine’ are too far apart in Hamming distance for one to evoke the other directly. However by attending the abstraction ‘container, which ignores all dimensions except ‘concave’, and ‘permeable’, the memetic mind decreases the apparent Hamming distance between them.
44. Under what conditions does that R increase faster than s? As it turns out, abstraction plays a crucial role. To determine how abstraction affects R, let us assume for the moment that memory is fully-connected. Clearly memory is not fully-connected, but this simplification illustrates some trends which also apply to a memory wherein sparseness is compensated for by restricted distributed activation. We will be conservative and limit the sort of retrieval event under consideration to abstraction, and the redescription of a meme as an instance of an abstraction (including analogical thought). Abstractions have n dimensions, where n ranges from a minimum of m to a maximum of M. RA, the number of ways a retrieval can occur through abstraction, equals the number of retrieval paths enabled by an n-dimensional abstraction, multiplied by the number of n-dimensional abstractions, summed over all values of n from m to M-1. The number of retrieval paths equals the number of memes that are instances of an n-dimensional abstraction = 2M-n. The number of n-dimensional abstractions is equal to the binomial coefficient of M and n. The result is multiplied by two, since an abstraction can evoke an instance, and likewise, an instance can evoke an abstraction.
|
RA |
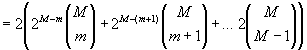 |
|
| (4) |
The key thing to note is that lower-dimensional memes enable exponentially more retrieval paths. Abstraction increases s by creating a new meme, but it increases R more, because the more abstract the concept, the greater the number of memes a short Hamming distance away (since |xi -ki| = 0 for the irrelevant dimensions). A second thing to note is that the number of abstractions at a given value of n increases up to M/2. Taken together, these points mean: the more deeply a mind delves into lower-dimensional abstractions, the more the distribution in FIGURE 1 rises and becomes skewed to the left. The effect is magnified by the fact that the more active a region of meme space the more likely an abstraction is to be positioned there, and thus abstractions beget abstractions recursively through positive feedback loops. Reminding incidents also contribute to R. Thus the more likely some meme will get activated and participate in a given retrieval. So, whereas R increases as abstraction makes relationships increasingly explicit, s levels off as new experiences have to be increasingly unusual in order to count as new and get stored in a new constellation of locations. Furthermore, when the carrying capacity of the memory is reached, s plateaus, but R does not. Thus, as long as the neuron activation threshold is large enough to permit abstraction and small enough to permit temporal continuity, the average value of n decreases. Sooner or later the system is expected to reach a critical critical percolation threshold such that R increases exponentially faster than s, as in FIGURE 4.
45. So long as R does indeed eventually increase faster than s, Groga’s memory becomes so densely packed that any meme that comes to occupy the focus is bound to be close enough in Hamming distance to some previously-stored meme(s) to evoke it. The memory (or some portion of it) is holographic in the sense that there is a pathway of associations from any one meme to any other. Together they form an autocatalytic set. What was once just a collection of isolated memories is now a structured network of concepts, instances, and relationships—a worldview. This most primitive level of autocatalytic closure is achieved when stored episodes are interconnected by way of abstractions just a few onionskin layers deep, and streams of thought zigzag amongst these superficial layers. A second level occurs when relationships amongst these abstractions are identified by higher-order abstractions at even deeper onionskin layers. Et cetera. Once Groga’s memory has defined an abstraction, identified its instances, and chunked them together in memory, she can manipulate the abstraction much as she would a concrete episode. Reflecting on an idea amounts to reflecting it back and forth off onionskin layers of varying depths, refining it in the context of its various interpretations. The conscious realization of the logical operators ‘and’, ‘or’ and ‘not’, are expected to significantly transform Groga’s worldview by enabling conscious symbol manipulation. Other particularly useful abstractions such as ‘mine’, ‘depth’, or ‘time’, as well as frames [Barsalou, in press], scripts [Schank & Abelson 1977], and schemas [Minsky 1985], are also expected to induce reorganization. Just as in a sand pile perched at the proverbial ‘edge of chaos’ a collision between two grains occasionally triggers a chain reaction that generates a large avalanche, one thought occasionally triggers a chain reaction of others that dramatically reconfigure the conceptual network. Rosch’s [1978] work on basic level categories suggests that the way we organize information is not arbitrary but emerges in such a way as to maximize explanatory power. It would not be surprising to find that the number of categories and their degree of abstraction exhibit the same kind of power law relationship as one finds in other emergent systems [Bak, Tang, & Weisenfeld 1988].
46. How does an interconnected worldview help Groga manifest the skills that differentiate a memetic mind from an episodic one? The capacity to maintain a stream of self-triggered memes enables her to plan a course of action, and to refine behavior by incorporating kinesthetic feedback into a meme sequence. The ability to generate abstractions opens up a vast number of new possibilities for Groga. It allows her to incorporate more of the structure of the world into her mental model of it. This increases behavioral flexibility by enabling her to define elements of the world in terms of their substitutable and complementary relationships. (For example, if she usually makes bows out of wood X, but she can’t find any wood X, and if wood Y is as strong and flexible as wood X, then wood Y might substitute for wood X.) The power of abstraction also enables her to express herself artistically by extricating memes from the constraints of their original domain and filtering the resulting pattern through the constraints of other domains. For example, she can translate the scene before her into a sequence of motor commands that render it as a cave painting or stone carving, or transform the pattern of information that encodes the sorrow she experienced at her child’s death into a song. Finally, abstraction enables Groga to communicate with others through spoken or nonverbal forms of language. This brings us to the issue: how does the worldview replicate?
47. Now that we have an autocatalytic network of memes, how does it self-replicate? In the OOL scenario, polymer molecules accumulate one by one until there are at least two copies of each, and their shell divides through budding to create a second replicant. In the OOC scenario, Groga shares concepts, ideas, stories, and experiences with her children and tribe members, spreading her worldview meme by meme. Categories she had to invent on her own are presented to and experienced by others much as any other episode. They are handed a shortcut to the category; they don’t have to engage in abstraction to obtain it.
48. Recall how the probability of autocatalysis in Kauffman’s simulation could be increased by raising either the probability of catalysis or the number of polymers (since it varied exponentially with M). Something similar happens here. Eventually, once enough of Groga’s abstractions have been assimilated, her tribe members’ memories become so densely packed that even if their neuron activation thresholds are higher than Groga’s, a version of Groga’s worldview snaps into place in their minds. Each version resides in a different body and encounters different experiences. These different selective pressures sculpt each ‘copy’ of Groga’s original worldview into a unique internal model of the world. Small differences are amplified through positive feedback, transforming the space of viable worldview niches. Individuals whose activation threshold is too small to achieve worldview closure are at a reproductive disadvantage, and over time eliminated from the population. Eventually the proclivity for an ongoing stream of thought becomes so firmly entrenched that it takes devoted yogis years of meditation to even briefly arrest it. There is selective pressure for parents who monitor their child’s progress in abstraction and interact with the child in ways that promote the formation of new abstractions the next level up. Recall the discussion in section II.4 concerning the incorporation of new polymer species by supracritical autocatalytic sets. This kind of parental guidance is analogous to handcrafting new polymers to be readily-integrated into a particular autocatalytic set; in effect it keeps the child’s mind perpetually poised at a supracritical state. Language provides a means for individuals to mutually enrich one another's worldviews, and to test their worldviews against each other, and in so doing prompt one another to penetrate deeper and deeper into the 'onion'.
49. Clearly social processes are an integral component of cultural evolution. In fact the origin of culture is often unquestioningly equated with onset of the capacity for social transmission. However, as many authors have pointed out [e.g. Darwin 1871; Plotkin 1988], although transmission is wide-spread in the animal kingdom, no other species has anything remotely approaching the complexity of human culture. Moreover, although in practice transmission plays an important role, is it crucial? If, for example, you were the only one human left on the planet, but you were able to live forever, would meme evolution grind to a halt? If you were to come up with some unique dance, wouldn’t you be exploring the space of possible dance memes even though no one was watching? If you found an ingenious way to fix a broken toaster, wouldn’t you still have invented a novel meme?
50. In biological evolution, transmission and replication go hand in hand; genetic information gets replicated and is transmitted to offspring. But that isn’t necessarily the only way of getting the job done. In memetic evolution, the most obvious means of meme replication is through social processes such as teaching or imitation, but there is a second form of replication that takes place within an individual. We noted earlier that in the mind of someone engaged in a stream of thought, each meme is a statistically similar variant of the one that preceded and prompted it. It is in this sense that they self-replicate without necessarily being transmitted to another host. Thus there need not necessarily be more than one individual for a meme to evolve. Nevertheless, although intra-individual meme replication is sufficient for evolving memes, the culture of a single individual would be extremely impoverished compared to that of a society of interacting individuals, because the number of memes increases exponentially as a function of the number of interacting memetic-level individuals. As a simple example, a single memetic individual who invents ten memes is stuck with just those ten memes. A society of ten interacting individuals, only one of whom has reached the memetic stage and can invent ten memes, is no better off; there are still just ten memes. In a society of ten non-interacting individuals, each of whom invents ten memes but doesn’t share them, each individual still has only ten memes. But in a society where each of the ten interacting individuals invents ten memes and shares them, each individual ends up with one hundred memes. The bottom line is: culture as we know it, with its explosive array of meaningful gestures, languages, and artifacts, depends on both intra-individual and inter-individual meme replication.
51. In fact it is possible that cognitive closure as described above first occured at the level of the group, within a collection of interacting individuals, and cognitive closure at the level of the individual came into existence some time later. (The two need not be mutually exclusive; it is possible that group-level closure could persist after the arrival of individual-level closure.)
52. In this section we explore some implications of the autocatalytic cognition hypothesis. This is the most speculative section of what is admittedly a speculative paper.
53. As noted in Section III.2, the penalty for having too low a neuron activation threshold is very high. Each meme has little relevance to the one that preceded it, and thinking is so garbled that survival tasks are not accomplished. On the other hand, too high a neuron activation threshold is not life-threatening. The focus is virtually always impacted with external stimuli or internal drives, and memory is reserved for recalling how some goal was accomplished in the past. This may be the situation present in most brains on this planet, and though not harmful, it has its own drawbacks. A stream of thought dies out long before it produces something creative. However, this may not be of practical consequence to other species. The advantages of a stream of thought would largely be lost on animals because they have neither the vocal apparatus nor the manual dexterity and freedom of upper limbs to implement creative ideas. (Language, for example, drastically increases the degrees of freedom of what can be expressed.) No matter how brilliant their thoughts were, it would be difficult to do something useful with them. Moreover, in an evolutionary line there is individual variation, so the lower the average activation threshold, the higher the fraction of individuals for which it is so low that they do not survive. It seems reasonable to suggest that animals are not prohibited from evolving complex cognition a priori, but that there is insufficient evolutionary pressure to tinker with the threshold until it achieves the requisite delicate balance to sustain a stream of thought, or to establish and refine the necessary feedback mechanisms to dynamically tune it to match to the degree of conceptual fluidity needed at any given instant. It may be that humans are the only species for which the benefits of this tinkering process have outweighed the risks.
54. Initially a child is expected to be unselective about meme acquisition, since (1) it doesn’t know much about the world yet, so it has no basis for choosing, and (2) its parents have lived long enough to reproduce, so they must be doing something right. However just as importing foreign plants can bring ecological disaster, the assimilation of a foreign meme can disrupt the established network of relationships amongst existing memes. Therefore the child develops mental censors that ward off internalization of potentially disruptive memes. Censors might also be erected when a meme is embarrassing or disturbing or threatening to the self-image [Minsky 1985]. This could be accomplished by temporarily increasing the activation threshold so as to prematurely terminate the meme’s assimilation into the worldview. Much as erecting a fence increases the probability that people will stay on either one side or the other, censorship warps the probability that a meme will partake in any particular stream of thought, such that the individual either avoids the censored meme or dwells on it excessively. (This seems to be consistent with our bipolar attitude toward highly censored subjects such as aggression and sexuality.) Thus censorship precludes incorporation of a meme into the autocatalytic portion of the memory, and thereby interferes with its holographic nature.
55. Categorization creates new lower-dimension memes, which makes the space denser, and increases susceptibility to the autocatalytic state. On the other hand, creating new memes by combining stored memes could interfere with the establishment of a sustained stream of thought by decreasing the modularity of the space, and thereby decreasing density. If indeed cross-category blending disrupts conceptual networking, one might expect it to be less evident in young children than in older children, and this expectation is born out experimentally [Karmiloff-Smith 1990]. There is evidence of an analogous shift in human history from an emphasis on ritual and memorization toward an emphasis on innovation [Donald, 1991]. As worldviews become more complex, the artifacts we put into the world become more complex, which necessitates even more complex worldviews, etcetera; thus a positive feedback cycle sets in.
56. We mentioned that animals are hardwired to respond appropriately to certain stimuli, as are humans. However, the ability of humans to develop worldviews with which they can make decisions about what action to take may obliviate the need for some of this hardwiring. Genetic mutations that interfere with certain regions of hardwiring may not be selected against, and may actually be selected for, because in the long run they promote the formation of concepts that generate the same responses but can be used in a more context-sensitive manner. However this increases the amount of computation necessary to achieve a workable worldview.
57. Returning briefly to the origin-of-life puzzle, recall that traditional attempts to explain how something as complex as a self-replicating entity could arise spontaneously entail the synchronization of a large number of vastly-improbable events. Proponents of such explanations argue that the improbability of the mechanisms they propose does not invalidate them, because it only had to happen once; as soon as there was one self-replicating molecule, the rest could be copied from this template. However, Kauffman’s theory that life arose through the self-organization of a set of autocatalytic polymers suggests that life might not be a fortunate chain of accidents but rather an expected event.
58. Although there is much evidence for this hypothesis, definitive proof that it is the correct explanation of how life originated will be hard to come by. However, if we are interested in the more general question of how information evolves, we now have another data point, another evolutionary process to figure into the picture. Culture, like biological life, is a system that evolves information through variation, selection and replication. In fact, it has two ‘layers’ of replication, one embedded in the other, and to actualize the inner layer of replication, every member of the culture must establish their own personal worldview, which generates their own unique autonomous stream of sequentially activated self-similar patterns. Consistent with Kauffman’s assertion that the bootstrapping of an evolutionary process is not an inherently improbable event, the ‘it only had to happen once’ argument does not hold water here because the cultural analog to the origin of life takes place in the brain of every young child. Autocatalysis may well be the key to the origin of not only biological evolution, but any information-evolving process.
59. Cultural evolution presents a puzzle analogous to the origin of life: the origin of an internal model of the world that both generates and is generated by streams of self-sustained, internally-driven thought. In this paper we explored a plausible scenario for how cultural evolution, like biological evolution, could have originated in a phase transition to a self-organized web of catalytic relations between patterns. TABLE 1 presents a summary of how the components of the proposed theory of cultural autocatalysis map onto their biological counterparts.
|
EVOLUTIONARY SYSTEM |
BIOLOGY |
CULTURE |
|
INFORMATION UNIT |
Polymer Molecule |
Meme |
|
INTERACTION |
Catalysis |
Reminding, retrieval, reconstruction |
|
AUTOCATALYTIC SET |
Catalytically closed set of polymer molecules = Primitive organism |
Network of inter-related memes = Worldview Worldv |
|
REPLICATION |
Duplication of each molecule, segregation via budding |
Correlation between consecutive memes / Social learning, teaching, imitation |
|
SELECTION |
Physical constraints on molecules, affordances and limitations of environment |
Associations, drives / Social pressures, affordances and limitations of environment |
|
VARIATION |
Novel food molecules, nonspecificity of catalysis, replication error |
Sensory novelty, blending / Expressive constraints, misunderstanding, et cetera |
TABLE 1: Components of an autocatalytic theory of biological evolution, and their cultural counterparts.
60. The scenario outlined here is nascent. Putting the pieces together would require the cooperation of neuroscientists, developmental psychologists, cognitive scientists, sociologists, anthropologists, archeologists, and perhaps others. Nevertheless, I know of no other serious attempt to provide a functional account of how memetic evolution got started. Whether or not the scenario outlined here turns out to be precisely correct, my hope is that it draws attention to the problem of cultural origins, suggests what a solution might look like, and provides a concrete example of how we gain a new perspective on cognition by viewing it as an architecture that has been sculpted to support a second evolutionary process, that of culture.
I would like to thank David Chalmers, Merlin Donald, Bruce Edmonds, Harold Edwards, Stuart Kauffman, Francis Heylighen, Norman Johnson, Wolfgang Klimesch, William Macready, and Mario Vaneechoutte for helpful discussion and comments on the manuscript.
1. Although this may not be completely accurate; see [Donald 1993] and accompanying commentary.
2. The magnitude of this number is perhaps better appreciated when we realize that it is equal to 10300.
3. In a reducing atmosphere there is no free oxygen present. The presence of ferrous (FeO) rather ferric (Fe2O3) iron in primitive rock leads us to believe that the earth's atmosphere was reducing when life began. (It isn't now.)
4. See [Kauffman 1993] for an interesting discussion of why error catastrophe becomes a serious problem as the parts of the system becomes more co-adapted.
5. The niche it filled still exists, so there is still selective pressure for it to evolve all over again. But the information has to 're-evolve' (as opposed to being retrieved from storage).The niche it filled still exists, so there is still selective pressure for it to evolve all over again. But the information has to 're-evolve' (as opposed to being retrieved from storage).
Bak, P., Tang, C., & Weisenfeld, K. (1988) Self-organized criticality. Phys. Rev. A 38: 364.
Barkow, J. H., Cosmides, L. & Tooby, J. (1992) The adapted mind: Evolutionary psychology and the generation of culture. Oxford University Press.
Barsalou, L. W. (In press) Perceptual symbol systems. Submitted to Behavioral and Brain Sciences.
Bickerton, D. (1990) Language and species, University of Chicago Press.
Cemin, S. C. & Smolin, L. (in press) Coevolution of membranes and channels: A possible step in the origin of life. Submitted to Journal of Theoretical Biology (October, 1997), adap-org/9709004.
Corballis, M. C. (1991) The lopsided ape: Evolution of the generative mind, Cambridge University Press.
Darwin, C. (1871) The descent of man, John Murray Publications.
Donald, M. (1991) Origins of the modern mind, Harvard University Press.
Donald, M. (1993a) Precis of Origins of the modern mind: Three stages in the evolution of culture and cognition. Behavioral and Brain Sciences 16: 737-791.
Donald, M. (1993b) Human cognitive evolution: What we were, what we are becoming. Social Research 60 (1): 143-170.
Eigen. M. & Schuster, P. (1979) The hypercycle: A principle of natural self-organization. Springer.
Farmer, J. D., Kauffman, S. A., & Packard, N. H. (1987) Autocatalytic replication of polymers. Physica D, 22 (50).
Hoyle, F. & Wickramasinghe, N. C. (1981) Evolution from space, Dent.
Joyce, G. F. (1987) Non-enzymatic, template-directed synthesis of informational macromolecules. In: Cold Spring Harbor Symposia on Quantitative Biology 52. Cold Spring Harbor Laboratory, New York.
Gabora, L. (1996a) Culture, evolution and computation. In Proceedings of the Second Online Workshop on Evolutionary Computation. Society of Fuzzy Theory and Systems. http://www.bioele.nuee.nagoya-u.ac.jp/wec2/papers/p023.html
Gabora, L. (1996b) A day in the life of a meme. Philosophica, 57, 901-938. Invited manuscript for special issue on concepts, representations, and dynamical systems. http://www.lycaeum.org/~sputnik/Memetics/day.life.txt
Gabora, L. (1997) The origin and evolution of culture and creativity. Journal of Memetics: Evolutionary Models of Information Transmission Vol. 1, Issue 1. http://www.cpm.mmu.ac.uk/jom-emit/1997/vol1/gabora_l.html
Hinton, G. E. & J. A. Anderson. (1981) Parallel models of associative memory. Lawrence Erlbaum Associates.
Kanerva, P. (1988) Sparse distributed memory, MIT Press.
Karmiloff-Smith, A. (1990) Constraints on representational change: Evidence from children’s drawing. Cognition 34: 57-83.
Karmiloff-Smith, A. (1992) Beyond modularity: A developmental perspective on cognitive science, MIT Press.
Kauffman, S. A. (1993) Origins of order, Oxford University Press.
Langton, C.G. (1992) Life at the edge of chaos. In: Artificial life II, eds. C. G. Langton, C. Taylor, J. D. Farmer & S. Rasmussen, Addison-Wesley.
Lee, D. H., Granja, J. R., Martinez J. A., Severin, K. & Ghadiri, M. R. (1996) A Self-Replicating Peptide. Nature. 382, 8 August, 525-528.
Lee, D. H., Severin, K., Yokobayashi & Ghadiri, M. R. (1997) Emergence of Symbiosis in Peptide Self-Replication through a Hypercyclic Network. Nature, 390, 11 December: 591-594.
Lieberman, P. (1991) Uniquely human: The evolution of speech, thought, and selfless behavior, Harvard University Press.
Miller, S. L. (1955) Production of some organic compounds under possible primitive earth conditions. Journal of the American Chemistry Society 77:23-51.
Minsky, M. (1985) The society of mind, Simon and Schuster.
Morowitz, H. J. (1992) The beginnings of cellular life, Yale University Press.
Olton, D. S. (1977) Spatial memory. Scientific American 236, 82-98.
Oparin, A. I. (1971) Routes for the origin of the first forms of life. Sub. Cell. Biochem. 1:75.
Orgel, L. E. (1987) Evolution of the genetic apparatus: A review. In Cold Spring Harbor Symposia on Quantitative Biology Vol. 52. Cold Spring Harbor Laboratory, New York.
Plotkin, H. C. (1988) The role of behavior in evolution, MIT Press.
Rumelhart, D. E. & J. L. McClelland. (1986) Parallel distributed processing: Explorations in the microstructure of cognition. MIT Press.
Rosch, E. (1987) Principles of categorization. In E. Rosch & B. B. Lloyd (Eds.) Cognition and categorization. Erlbaum.
Schank, R., & Abelson, R. P. (1977) Scripts, plans, goals, and understanding: An inquiry into human knowledge structures. Lawrence Erlbaum Associates.
Severin, K., Lee, D. H., Kennan, A. J. & Ghadiri, M. R. (1997) A Synthetic Peptide Ligase. Nature, 389, 16 October, 706-709.
Tomasello, M., Kruger, A. C. & Ratner, H. H. (1993) Cultural learning. Behavioral and Brain Sciences, 16, 495-552.
Tooby, J. & Cosmides, L. (1989) Evolutionary psychology and the generation of culture, Part I. Ethology and Sociobiology, 10, 29-49.
Weisberg, R.W. (1986) Creativity: Genius and other myths, Freeman.